
Условия укрепления и ослабления связей
Системная нейрофизиология (nan)
|
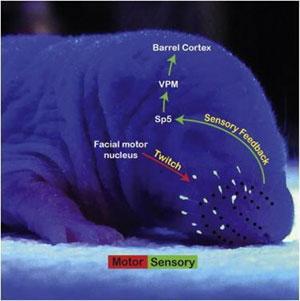
В ходе индивидуального развития организма сила передачи сигнала в отдельно взятом синапсе (контакте между двумя нейронами) может как увеличиваться (синаптическая потенциация), так и уменьшаться (синаптическая депрессия). Синаптическая депрессия в ряде случаев служит инструментом ослабления и в конечном счете устранения ненужных нейронных контактов. Неожиданный по своему молекулярному механизму пример синаптической депрессии, основанный не на времени прихода сигналов на синапс, а на их количестве и группировке (паттерне), был обнаружен в нейронах сенсорной коры молодых мышей, воспринимающей и обрабатывающей информацию от органов осязания — вибрисс («усов»).
Сведения об окружающем мире животные получают от органов чувств. Доля информации, поступающей от отдельных сенсорных систем (слуха, зрения, обоняния, осязания и т.д.), меняется в течение всей жизни, причем наиболее радикальные и быстрые изменения происходят до взросления организма. На уровне отдельных нервных клеток изменение доли информации от различных сенсорных систем проявляется в усилении или ослаблении связей между нейронами — синапсов, — обеспечивающих проведение импульсов от органов чувств в центральную нервную систему и дальнейшую обработку этих импульсов в мозге.
Мыши, как и другие грызуны, растут очень быстро. На 14-й день мышата открывают глаза, а на 21-й становятся полностью самостоятельными. Получается, что до начала третьей недели жизни мыши узнают об окружающем их мире почти исключительно за счет осязания и обоняния, а затем у них есть еще около семи дней на то, чтобы «отдать часть управления» зрительной системе. То есть какие-то связи между нейронами, раньше получавшими информацию о прикосновении и запахе, должны быть ослаблены или вовсе прерваны.
Значительную часть осязательной (тактильной) информацию мыши получают от вибрисс («усов»). После вибрисс нервные пути, несущие тактильную информацию, расходятся. Тот путь (цепочка нейронов), который нас интересует проходит от основания вибрисс (фолликулов) через ядро тройничного нерва в вентральное постеромедиальное ядро (VPM на рис. 1) , где сходится тактильная информация и от других областей тела. Далее информация от вибрисс из таламуса попадает в (S1), в ту её область, которая получает сигналы исключительно от «усов» (см. рис. 1). Как и другие зоны новой коры () больших полушарий, S1 имеет слоистое строение (состоит из шесть слоёв); сигналы от таламуса получает четвёртый слой коры.
Точность поступающих сигналов обеспечивается особым строением четвёртого слоя, при котором каждой вибриссе соответствует определённая группа клеток. Все нейроны внутри такого ансамбля, называемого «бочонками соматосенсорной коры», получают информацию только от одного «уса». На окрашенных срезах неокортекса обсуждаемые группы клеток выглядят как последовательные ряды (рис. 2).
< Блок иллюстрации >
|
Нейроны бочонков получают сигналы от таламуса, а передают их, в свою очередь, клеткам соматосенсорной коры. Этот-то контакт (в дальнейшем будем называть его «синапс 4–2/3» по номеру слоя неокортекса) и был выбран для исследования. Причина такого выбора — высокая пластичность синапсов 4–2/3 на третьей неделе жизни мыши: обнаружили, что у мышат в возрасте 15–21 день может происходить ослабление (долговременная депрессия) некоторых связей нейронов слоёв 4 и 2/3. Однако механизм этой депрессии понятен не был.
Вспомним, как устроен химический синапс — место контакта двух нейронов, разделённых синаптической щелью. Оба нейрона могут менять потенциал на своих мембранах (деполяризоваться или гиперполяризоваться). Изменения потенциала передаются через синаптическую щель в виде молекул нейромедиаторов (переносчиков нервного сигнала) — веществ, выделяемых нейроном и служащих для изменения потенциала воспринимающей их клетки. Один из нейронов в синапсе всегда выполняет роль подающего сигналы и называется пресинаптическим, то есть сигнал через него проходит раньше, чем нейромедиатор через синпатическую щель. Второй нейрон получает информацию от первого и называется постсинаптическим, так как информация проходит через него позже, чем медиатор через синаптическую щель. Сила связи между двумя нейронами, выраженная в амплитуде изменения мембранного потенциала одного из них или в количестве выделяемого нейромедиатора, может изменяться. Такое изменение называют синаптической пластичностью.
Первое, что требовалось выяснить, — как на клеточном уровне выражается тактильное воздействие на отдельную вибриссу живой мыши. Для этого учёные регистрировали электрическую активность нейронов четвёртого слоя коры, возникающую в ответ на прикосновение к одной вибриссе.
Нейроны обладают свойствами возбудимости и раздражимости. Это означает, что в ответ на отклонение «уса» нервная клетка, с ним связанная, проявит электрическую активность: разность потенциалов на её мембране, будучи отрицательной, изменится, приблизившись к нулю и, преодолев некоторый предел своего изменения (порог генерации потенциала действия, ПД), резко вырастет, став на миллисекунды положительной (произойдёт деполяризация мембраны), и затем так же резко вернётся к своему значению до возбуждения. Пресинаптическая нервная клетка генерирует ПД, после чего постсинаптическая клетка тоже может сгенерировать ПД и передать сигнал дальше по цепи. Если вторая клетка в ответ на сигнал от первой не сгенерирует ПД, сигнал дальше не пойдёт.
Выяснилось, что каждая клетка бочонка реагирует на такое прикосновение одним-четырьмя потенциалами действия в течение 50–200 миллисекунд после тактильной стимуляции вибриссы.
Далее надо было понять, какая именно активность нейрона четвёртого слоя может привести к ослаблению его связи с клеткой второго-третьего слоёв. На этом этапе исследователи работали уже не на живых мышах, а на срезах соматосенсорной коры. В каждом таком срезе регистрировали активность попарно связанных друг с другом нейронов синапса 4–2/3 методом пэтч-клэмп. В каждой такой паре клетку слоя 4, играющую роль пресинаптической, деполяризовали так, чтобы она с определённой частотой генерировала потенциал действия (ПД). Клетка слоя 2/3, постсинаптическая в этой паре, отвечала на пришедший от пресинаптического нейрона сигнал возбуждающим постсинаптическим потенциалом (ВПСП). Величина ВПСП в ответ на один ПД, служивший тестирующим стимулом, в ходе часового эксперимента практически не изменялась. Изменение скорости достижения максимальной амплитуды ВПСП и величины этого потенциала происходило после серии из 100 стимуляций, точно копировавших варианты активности нейронов четвёртого слоя, записанных в ходе экспериментов на живых мышатах (рис. 3).
< Блок иллюстрации >
|
Оказалось, что ответ постсинаптической клетки синапса 4 2/3 усиливается, если потенциал действия на ней возникает после прихода аналогичного сигнала с пресинаптического нейрона (рис. 3, A), и ослабевает, если сначала возбуждается постсинаптическая клетка, а пресинаптическая разряжается после (рис. 3, B). Такие потенциация (усиление) и депрессия (ослабление) связей вполне объяснимы: синапс, условно говоря, правильный, если информация сначала поступает на его вход, а потом уже идёт на выход, но не наоборот.
Но как нервная клетка выясняет, какие из связей ей стоит поддерживать, а какие — нет? Любой нейрон может иметь несколько связей с несколькими другими нейронами, причем активность каждой связи не зависит от активности других. Если от пресинаптического (для данного синапса) нейрона много раз подряд (или долгое время с определённой периодичностью) на постсинаптический нейрон приходит сигнал, способный вызвать в нём ПД, то связь между этими двумя нейронами усиливается. Это явление называется . (Здесь важно неоднократное повторение импульсов от пресинаптического нейрона и то, что потенциалы действия в принимающем, постсинаптическом, нейроне появляются позже, чем в пресинаптическом.) Механизмы усиления связей между нейронами изучены гораздо более тщательно, чем процессы, ведущие к их ослаблению. Долгое время был известен только один механизм ослабления связей нейронов — прерывание связи с клеткой, «не успевающей» вовремя подать сигнал. Допустим, пресинаптический нейрон подаёт сигнал на принимающий нейрон, когда тот уже возбудился по каким-то другим причинам (например, нейрон из другой нервной цепи подал на него сигнал раньше, чем «наш» нейрон, а «наш» — «опоздал»). В таком случае постсинаптическая нервная клетка ослабляет связь с «опаздывающей» клеткой.
Однако в обсуждаемом исследовании ослабление синаптической передачи было ещё сильнее, если авторы статьи искусственно не давали принимающей клетке реагировать на ПД, приходящий от пресинапса (рис. 3, C). Этот факт позволил исследователям предположить, что механизм депрессии синапса 4 2/3 отличается от классического Хеббовского. Но как именно?
Если порядок возбуждения пре- и постсинаптического нейрона не имеют значения, то, вероятно, важны количество и частота следования (группировка) потенциалов действия, приходящих от клетки слоя 4 (получается своего рода азбука Морзе). Другими словами, существует определённая «фраза» (её называют паттерном), выдаваемая пресинаптической клеткой, на которую постсинаптическая должна ответить потенциалом действия, а в случае игнорирования такой «фразы» синаптическая связь ослабится как неэффективная или ненужная.
Чтобы понять, как выглядит такой сигнал на языке ПД, исследователи деполяризовали нейрон слоя 4 так, чтобы он разряжался двумя-четырьмя спайками (нервными импульсами), идущими либо через равные промежутки времени, либо сгруппированными по несколько ПД. Все варианты группировки потенциалов действия (паттерны) были взяты из опытов на живых мышах (см. выше).
Оказалось, что наиболее сильную депрессию синаптической связи нейронов слоёв 4 и 2/3 вызывает игнорирование паттерна 3+1: пресинапс разряжается три раза подряд, после чего через промежуток времени, примерно равный длительности этих трёх спайков (один спайк соответствует одному пику на рисунке), «фраза» завершается последним потенциалом действия. Новый тип долговременной депрессии, вызванный отсутствием ответа постсинаптической клетки на такой сигнал, назвали паттерн-зависимым.
Чтобы выяснить молекулярный механизм паттерн-зависимой долговременной депрессии (ДВД), авторы вводилиблокаторы тех или иных рецепторов (комплексов белковых молекул, служащих для распознавания сигнала, принесённого к нейрону в форме нейромедиатора) или ферментов в одну из клеток синапса 4–2/3 или же в межклеточное пространство, в зависимости от мишени и механизма действия конкретного вещества. При выборе блокаторов учёные исходили из имеющейся информации о классическом механизме ДВД.
Известно, что в случае Хеббовского варианта депрессии основную роль в ослаблении контакта двух нейронов играет постсинаптическая клетка. Последовательность биохимических реакций (молекулярный каскад) в ней включает повышение внутриклеточной концентрации кальция, который вызывает выброс нетипичных нейромедиаторов — эндогенных каннабиноидов. Последние отличаются тем, что выделяются не пресинаптическим, а постсинаптическим нейроном и действуют на рецепторы, расположенные на клетках нейроглии, — каннабиноидные рецепторы CB1. В ответ на активацию CB1 в клетке глии также повышается концентрация кальция, и она выделяет нейромедиатор глутамат, основной переносчик сигнала в данном синапсе. Нейроглиальный глутамат активирует пресинаптические NMDA-рецепторы (ионотропные рецепторы глутамата), что, в свою очередь, тормозит дальнейший выброс медиатора пресинапсом.
Авторы обсуждаемой статьи последовательно блокировали активацию постсинаптических NMDA-рецепторов, повышение внутриклеточной концентрации ионов кальция в той же клетке, эндоканнабиноидные рецепторы нейроглии и активность глиальных элементов в целом. Ни одно из этих воздействий не повлияло ни на возникновение, ни на поддержание ДВД. Зато дезактивация пресинаптических NMDA-рецепторов, а также кальций-зависимого фермента (CN), задействованного и в долговременной депрессии «классического» типа, блокировала появление паттерн-зависимой ДВД в рассматриваемом примере.
На основе полученных результатов учёные составили сравнительную схему молекулярных механизмов Хеббовской и паттерн-зависимой форм ДВД (см. рис. 4).
< Блок иллюстрации >
|
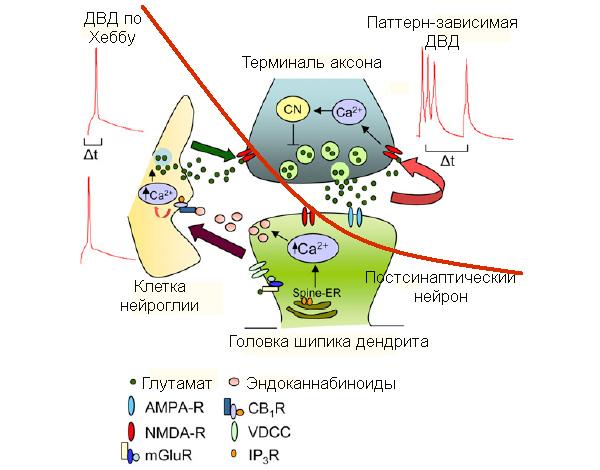
Схема наглядно демонстрирует, что постсинаптическая клетка в паттерн-зависимой ДВД фактически не задействована: все механизмы депрессии такого типа завязаны на пресинапсе, и роль постсинапса здесь сводится к тому, чтобы не отвечать на приходящие на него сигналы. Это первый зарегистрированный случай подобного ослабления синаптических связей. Однако полной и подробной картины того, что происходит в пресинаптической клетке во время проявления такого рода ДВД пока что нет. Поэтому первоочерёдная задача исследователей — прояснить точные каскады событий внутри пресинаптического нейрона при паттерн-зависимой депрессии.
Источник: Antonio Rodríguez-Moreno, Ana González-Rueda, Abhishek Banerjee, A. Louise Upton, Michael T. Craig, Ole Paulsen. Presynaptic Self-Depression at Developing Neocortical Synapses [http://www.cell.com/neuron/fulltext/S0896-6273(12)00956-7] // Neuron. 9 January 2013. V. 77. Issue 1. P. 35–42. (Статья находится в свободном доступе.)
См. также:
1) Jesper Sjöström, Wulfram Gerstner. // Scholarpedia, 5(2):1362. (2010).
2) Natalia Caporale, Yang Dan. // Annual Review of Neuroscience. V. 31. P. 25–46. (July 2008).
3) .
4) , «Элементы», 22.06.2010.
Светлана Ястребова
От Fornit:
Стоит иметь в виду, что при формировании функции распознавания важно закреплять связи всех участников с их постсинаптическим партнером, который и станет затем распознавателем данного профиля. В этом случае принцип Хебба для вторичных и третичных зон должен реализоваться с некоторым безразличием к неодновременностям входных возбуждений (в виду того, что они приходят от разных рецепторов с разными задержками).
Источник:
Дата создания: 18.03.2013
Последнее редактирование: 18.03.2013
Относится к аксиоматике: Системная нейрофизиология.
Другие страницы раздела "Условия образования связей":
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя
Схема нервных путей, обеспечивающих передачу сенсорного сигнала от вибрисс в кору головного мозга. Изображение из статьи