
Усиление контраста латеральным торможением
Системная нейрофизиология (nan)
Схема сенсорной системы на рис. 8.7 сильно упрощена, поскольку не отражает дивергенции и конвергенции, обсуждавшихся в предыдущем разделе. «Молнии» изображают два точечных стимула, находящиеся на небольшом расстоянии друг от друга и действующие на группу рецепторов одновременно. Результирующее распределение возбуждения таково, что стимулы первоначально не различаются
ГЛАВА 8. ОБЩАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ 187
как отдельные раздражители. Однако за счет подбора параметров латерального торможения удается достичь их полного разделения через два этапа синаптической передачи.
Чтобы смоделировать латеральное торможение, первичные афференты на схеме на рис. 8.7 соединяются со следующими вторичными нейронами не только непосредственно. Каждый из них возбуждает своими аксонными коллатералями интернейроны (черные кружки на рисунке), а каждый из последних тормозит соседний вторичный нейрон (белые кружки). Степень торможения, естественно, зависит от степени возбуждения интернейронов, а значит, и первичных афферентов. Модель предполагает, что каждому синапсу между первичным и вторичным нейронами соответствует коэффициент усиления, равный 3; при этом торможение посредством интернейронов (один возбуждающий и один тормозный синапс, расположенные последовательно) дает общий коэффициент усиления — 1. Как обычно в случае постсинаптического торможения, тормозные влияния (потенциалы) вычитаются из возбуждающих. На следующем этапе синаптической передачи этот процесс повторяется с теми же коэффициентами передачи. На рис. 8.7 уровни возбуждения и торможения указаны цифрами. Возникающее в результате чистое возбуждение можно понимать, например, как «число потенциалов действия» (в красных рамках—гипотетические записи импулъсации аксонов). Поэтапные изменения легко вычислить с учетом приведенных выше допущений.
Выделение сложных признаков высшими сенсорными нейронами. Усиление контраста не единственный способ выделения информации высшими сенсорными нейронами. В проекционных и ассоциативных областях коры головного мозга отдельные нейроны выделяют из сенсорного возбуждения гораздо более сложную информацию. Например, в соматосенсорной области есть клетки, кодирующие скорость и направление движения раздражителя по коже. Зрительная кора содержит простые и сложные клетки (см. с. 258), выделяющие отдельные признаки геометрии и движения зрительных стимулов. Более подробно организация различных сенсорных проекционных зон коры обсуждается в главах, посвященных соответствующим сенсорным системам. Здесь, говоря об общей сенсорной физиологии, важно подчеркнуть, что наши центральные сенсорные системы, особенно корковые, анализируют входные сигналы, обеспечивая их сознательное восприятие, путем выделения, или абстрагирования, сенсорной информации.
8.3. Общая субъективная сенсорная физиология
Основные характеристики ощущений
В начале этой главы субъективная сенсорная физиология определялась как область исследований, занимающаяся соотношением между физикохимическим миром стимулов и субъективным миром ощущений и восприятий.
По традиции у ощущений различают четыре основных характеристики - интенсивность, качественный тип. временную длительность и пространственную протяженность. Для некоторых сенсорных модальностей в последнем случае лучше говорить о местоположении. Если обратиться к слуху или обонянию, это означает способность обнаружить (локализовать) источник звука или запаха. В случае вкуса, например, пространственной характеристики вообще нет.
Прежде всего рассмотрим первые два из перечисленных параметров. Начнем с качественного типа. Дело в том, что ощущения принципиально различны по своей природе. «Видение»-это нечто совершенно иное, нежели «слышание». Зрение, слух, обоняние, вкус и т.д. называют сенсорными модальностями. Каждая из них включает различные качественные типы ощущений. Красный цвет - один тип зрительных ощущений, зеленый - другой.
В классической медицине различали пять «чувств»: зрение, слух, вкус, обоняние и осязание. Теперь выделяют несколько дополнительных модальностей (например, чувства температуры и равновесия). Вопрос о количестве присущих человеку «чувств» всегда будет связан с особенностями интерпретации этого понятия.
Следует также иметь в виду, что некоторые виды позвоночных обладают сенсорными органами, которых у нас нет. Например, лицевые ямки некоторых змей содержат рецепторы инфракрасного излучения, позволяющие обнаруживать теплокровную добычу. У некоторых рыб есть органы, чувствительные к электрическим полям. Они улавливают токи, связанные с мышечной активностью жертвы, которая зарылась в донный песок, и помогают ее локализовать. Современная техника позволяет сконструировать аналогичные по свойствам приборы, однако, чтобы мы могли ими пользоваться, информация на выходе должна быть преобразована в зрительные или слуховые сигналы.
Все пять классических чувств человека относятся к экстероцепции (см. с. 181). О существовании проприоцепторов и интероцепторов не было известно, пока физиологи их не открыли.
Особые трудности возникают при классификации боли и других неприятных ощущений, например зуда. Боль-это сенсорная модальность, а зуд, возможно, следует считать ее качественным типом. Однако она отличается от других чувств тем, что присутствует во всех категориях ощущенийинтеро-, проприо- и экстероцепции. Ноцицепторы, воспринимающие боль, занимают среди кожных рецепторов (все это экстероцепторы) особое положение, поскольку их информация касается не внешнего мира, а повреждений собственного тела. Иными словами, боль как модальность связана с телом. Подробнее физиология боли рассмотрена в гл. 10.
Именно для качественной характеристики ощущений Иоганнес Мюллер (1837) сформулировал
188 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
закон специфических сенсорных энергий, упоминавшийся выше: модальность зависит не от стимула, а от раздражаемого им органа (см. с. 180).
Сенсорные модальности нельзя сопоставлять друг с другом с помощью прямых измерений. Однако на уровне качественных типов ощущений такие измерения вполне возможны. Если медленно изменять частоту тона, будет обнаружен некоторый порог, т.е. различие в частотах, достаточное для заметного на слух перехода к другому качеству звучания. Точно так же, меняя частоту электромагнитного излучения, можно определить порог воспринимаемого изменения цвета.
Интенсивность ощущения: абсолютные и дифференциальные пороги
Интенсивность ощущений наиболее подробно изучалась психофизическими методами. Интерес к тому, можно ли измерить эту характеристику, привел Г. Фехнера к разработке примерно в 1850 г. первой научной методики количественного описания субъективного опыта. Он же вывел первый психофизический закон, устанавливающий количественную связь между физической интенсивностью (φ) и силой ощущения (ψ). До этого господствовала доктрина Декарта (1596-1650), отрицавшего возможность измерений в субъективной сфере («res cogitans»). Центральным понятием психофизики стало понятие сенсорного порога.
Абсолютный порог определяется как наименьший по интенсивности стимул, способный вызвать определенное ощущение. Некоторые авторы обычно ограничивают смысл этого термина, понимая под ним наиболее низкий порог, достижимый при оптимальных условиях стимуляции и адаптации. Например, пороговые значения для слуха зависят от частоты звука (с. 287), а для зрения — от времени адаптации (с. 255).
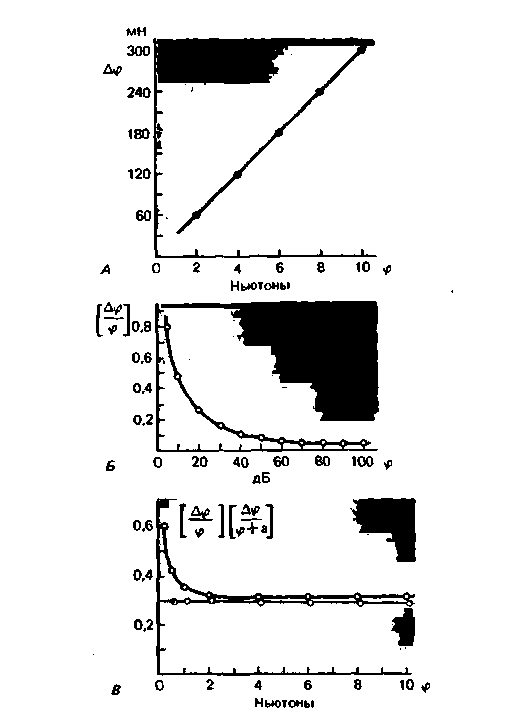
Дифференциальные пороги. В надпороговом диапазоне определяют еще один вид порога-«едва заметное различие». Это величина, на которую один стимул должен отличаться от другого, чтобы их разница воспринималась человеком. Э. Вебер (1834), экспериментируя с ощущением силы, первым показал, что для различения веса двух предметов их разница должна быть больше, если оба они тяжелые, и меньше, когда оба легкие. На рис. 8.8, А показано соотношение между предъявляемым весом и его едва заметным увеличением. В среднем диапазоне такой стимуляции зависимость линейная; иными словами, чтобы превысить дифференциальный порог, требуется одинаковая часть начального веса. Минимальное различимое изменение интенсивности стимуляции (Δφ) составляет постоянную
долю (с) ее исходной интенсивности (φ). Это закон Вебера, выражаемый уравнением
![]() (1)
(1)
Значит, ![]() -константа для всех интенсивностей
-константа для всех интенсивностей
стимула. Данное правило, действительно, выполняется в широком их диапазоне для многих сенсорных модальностей. Однако по мере приближения к абсолютному порогу коэффициент Вебера (Δφ/φ) обычно растет. Это проиллюстрировано на рис. 8.8, Б для громкости тона. Очевидно, в этом случае закон Вебера начинает действовать только при интенсивности стимула на 40 дБ выше абсолютного порога: начиная с этого уровня, коэффициент Вебера остается постоянным. Аналогичные кривые получены и для других модальностей.
Коэффициент Вебера-полезная мера относительной чувствительности сенсорных систем. Нельзя математически сравнить чувствительность глаза к силе света с чувствительностью уха к уровню звукового давления, но можно сопоставить между собой безразмерные коэффициенты Вебера для этих модальностей. В подобных опытах глаз несколько лучше, чем ухо, улавливает разницу адекватных стимулов по интенсивности.
Для соответствия экспериментальным данным закон Вебера можно переформулировать следующим образом:
![]() (2)
(2)
где а - константа, обычно небольшая по численному значению. Когда интенсивность стимула (φ) достаточно велика, членом «а» можно пренебречь, и выражение, соответствующее исходной форме закона Вебера, описывает наблюдения достаточно точно (рис. 8.8,5).
В некотором смысле константа «а»-просто поправочный член, обеспечивающий более точное соответствие экспериментальным данным. Однако его можно рассматривать и как «шум» в сенсорном канале. С точки зрения нейрофизиологии такой «шум» объясняется спонтанной активностью сенсорных нейронов, особенно высших уровней, т. е. их импулъсацией в отсутствие действующих на сенсорный орган стимулов. Вероятно, она добавляется к активности нейронов в ответ на стимуляцию, поэтому также влияет на порог, необходимый для едва заметного увеличения сигнала в ЦНС. Спонтанная активность мала по сравнению с реакцией на сильные стимулы, однако, когда стимул слаб, влияет на величину (![]() ).
).
В соответствии с этой нейрофизиологической интерпретацией, абсолютный порог должен быть равен минимальной интенсивности стимула, достаточной для возбуждения сенсорной системы, разли-
ГЛАВА 8. ОБЩАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ 189
|
Рис. 8.8. Коэффициент и закон Вебера. А. Соотношение между величиной раздражения (φ) и его увеличением (Δφ), необходимым для того, чтобы превзойти дифференциальный порог ощущения силы. Б. Зависимость коэффициента Вебера ( |
чимо большего спонтанной активности. Данный подход используется в некоторых психофизических теориях (например, в рассматриваемой ниже «сенсорной теории принятия решений»), В разделе, посвященном интегративной сенсорной физиологии, мы вернемся к вопросу о том, действительно ли спонтанная активность решающим образом влияет на абсолютный порог.
Фехнеровская психофизика
Помимо определения порогов, можно ли как-то еще измерить субъективные ощущения? Фехнер предложил использовать дифференциальные пороги
|
Рис. 8.9. Схематическое изображение закона Фехнера. По оси абсцисс интенсивность стимула, по оси ординат- дифференциальные пороги |
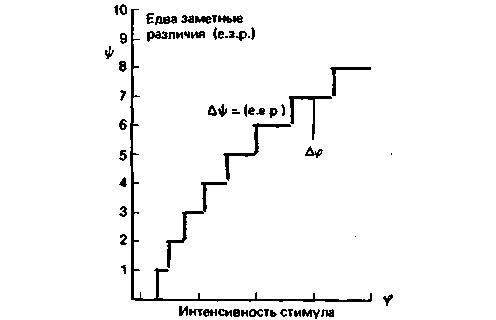
для построения шкалы интенсивности ощущений (![]() ). Нуль на ней соответствует абсолютному порогу, следующее ощущение больше точно на величину одного едва заметного различия, следующее-еще на одну такую величину и т.д. Поскольку каждая ступенька соответствует наименьшему возможному усилению ощущения, этот прирост и берется за основную единицу интенсивности ощущения. На рис. 8.9 показан график зависимости интенсивности ощущения от интенсивности стимула. Это соотношение между ψ и φ описывается уравнением, известным как психофизический закон Фехнера. Поскольку стимул увеличивается путем последовательного прибавления все возрастающей величины Δφ, получается логарифмическая кривая (рис. 8.9). Отсюда закон утверждает: при линейном увеличении интенсивности ощущения (ψ) интенсивность стимула (φ) растет логарифмически. Если психофизическую функцию (рис. 8.9) представить в линейно-логарифмической системе координат, кривая превратится в прямую (рис. 8.13). Фехнеровский закон выражается уравнением
). Нуль на ней соответствует абсолютному порогу, следующее ощущение больше точно на величину одного едва заметного различия, следующее-еще на одну такую величину и т.д. Поскольку каждая ступенька соответствует наименьшему возможному усилению ощущения, этот прирост и берется за основную единицу интенсивности ощущения. На рис. 8.9 показан график зависимости интенсивности ощущения от интенсивности стимула. Это соотношение между ψ и φ описывается уравнением, известным как психофизический закон Фехнера. Поскольку стимул увеличивается путем последовательного прибавления все возрастающей величины Δφ, получается логарифмическая кривая (рис. 8.9). Отсюда закон утверждает: при линейном увеличении интенсивности ощущения (ψ) интенсивность стимула (φ) растет логарифмически. Если психофизическую функцию (рис. 8.9) представить в линейно-логарифмической системе координат, кривая превратится в прямую (рис. 8.13). Фехнеровский закон выражается уравнением
![]() (3)
(3)
где ![]() -интенсивность ощущения, k - константа,
-интенсивность ощущения, k - константа, ![]() -интенсивность стимула и
-интенсивность стимула и ![]() - его абсолютный порог. Данный психофизический закон основан на двух главных допущениях.
- его абсолютный порог. Данный психофизический закон основан на двух главных допущениях.
Он действует при справедливости закона Вебера. При очень слабой стимуляции веберовский коэффициент не постоянен. Значит, закон Фехнера в этой области не действует.
Допускается также, что все приросты интенсивности ощущения равны, т. е. что пороговое изменение слабого и сильного стимулов ощущается одинаково. Рассматривая закон Стивенса, мы убедимся в некорректности данного допущения; в результате справедливость «закона» Фехнера сильно ограниче-
190 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
на. В свете этих данных лучше было бы говорить о психофизическом соотношении Фехнера.
Заслуга Фехнера в том. что он первым ввел пригодную для использования шкалу «интенсивности ощущения». Он вполне заслужил титул отца экспериментальной психологии. Теперь мы знаем, что ордината ψ в законе Фехнера выражает скорее разрешающую способность, чем интенсивность ощущения. Ограничения этой шкалы обсуждаются ниже.
Соотношение Фехнера использовалось для выведения логарифмических систем мер, применимых в сенсорной физиологии, например шкалы децибелов и фонов (см. с. 287). В промежуточном диапазоне частот н интенсивностей звуков минимальное заметное различие соответствует примерно 1 дБ.
Методы определения порогов
В предыдущем разделе было показано, что абсолютные и дифференциальные пороги относятся к фундаментальным психофизическим понятиям. Теперь вернемся к вопросу о том, как их измерять. Примеры относятся к абсолютным порогам, но применимы и к дифференциальным.
Реакции биологических систем неустойчивы, поэтому при тестировании слабыми стимулами испытуемый иногда их воспринимает, иногда-нет. Следовательно, нельзя определить порог как интенсивность стимула, ниже которой он никогда не ощущается, а выше-всегда заметен. Стимул нужно предъявлять несколько раз, после чего, обработав результаты статистически, оценить «истинный», средний порог. Есть несколько методик определения порога, некоторые из которых разработаны еще Фехнером.
К классическим относится метод пределов. Исходный стимул здесь далек от предполагаемого порогового значения. Например, интенсивность сначала устанавливают такой высокой, что испытуемый легко воспринимает стимул, а затем снижают до тех пор, пока она не окажется подпороговой. Затем тестирование начинают снова-на этот раз с очень слабых стимулов, которые усиливают до достижения порога. Такие восходящие и нисходящие последовательности повторяют несколько раз и берут среднее значение полученных пороговых величин. Интенсивность стимула можно записывать автоматически на миллиметровой бумаге, так что для выявления колебаний порога во времени необходимо будет лишь провести линию, соединяющую концы отрезков получаемых кривых.
Субъективная сенсорная физиология и поведенческие эксперименты на животных. Метод пределов, как и другие способы измерения порогов, можно использовать и в экспериментах на животных.
|
Рис. 8.10. Измерение зрительного порога голуоя в поведенческом эксперименте. А. Схема экспериментальной установки. Голубь клюет ключ А, если видит свет, в результате чего сила света уменьшается. Клевание ключа Б, когда света не видно, увеличивает интенсивность последующего светового стимула. Б. Постепенное снижение порога после выключения яркого фонового освещения. Получена кривая темновой адаптации голубя (по [8, 9] с изменениями) |
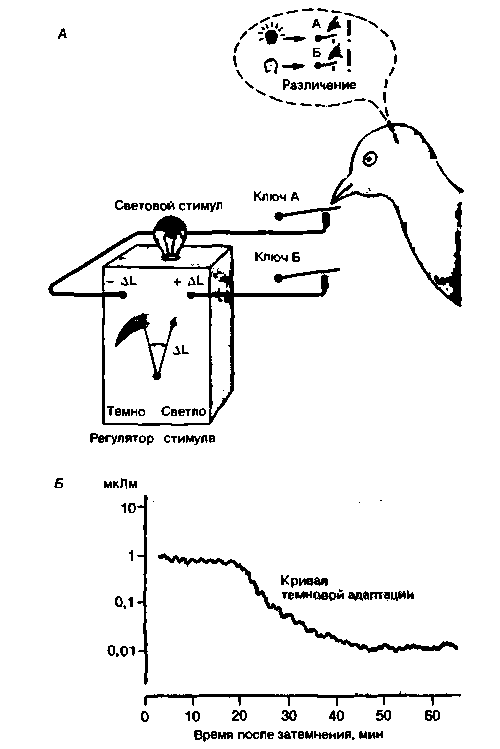
Пример показан на рис. 8.10. У голубя выработан условный рефлекс (см. гл. 6) получать пищевое вознаграждение после клевания ключа А, если он видит свет, и ключа Б, если света не замечает. При этом ключ А снижает интенсивность светового стимула, а ключ Б увеличивает ее. В ходе такого поведенческого эксперимента получаются восходящие и нисходящие кривые силы света, точки перегиба на которых соответствуют зрительному порогу. Если клетку с голубем изолировать от света, то непрерывное измерение порога таким способом даст кривую темновой адаптации (см. с. 255), сходную с получаемой для человека. Этот пример
ГЛАВА 8. ОБЩАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ 191
ясно показывает, что методы субъективной сенсорной физиологии применимы и в поведенческих экспериментах на животных.
Подобные эксперименты упоминаются здесь в разделе о субъективной сенсорной физиологии, поскольку своими методами они очень сходны с психофизическими опытами на людях. Можно, естественно, считать, что измеренные параметры поведения отражают субъективные ощущения животных, которые вовсе не обязательно сравнимы с сознательными переживаниями человека.
В одной из модификаций метода пределов испытуемый сам управляет стимуляцией (например, поворачивая рукоятку), усиливая ее до тех пор, пока она не станет ощущаться, или ослабляя до невоспринимаемого уровня. Преимущество данного метода настройки в более активной роли испытуемого по сравнению с другими психофизическими процедурами, при которых неизбежна усталость, вызываемая монотонностью эксперимента. Недостаток его-в невозможности выдержать одно и то же время стимуляции.
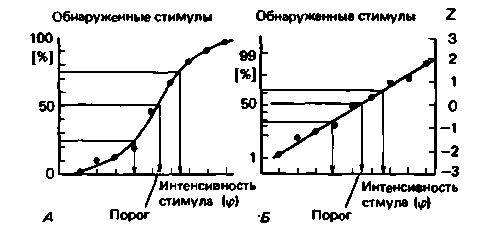
Психометрическая функция. Гораздо сложнее процедура предъявления различных по интенсивности стимулов в случайном порядке. Однако она позволяет получить более точные данные. Это так называемый метод постоянных стимулов. Они подбираются таким образом, что самый слабый почти никогда не замечается, а самый сильный ощущается почти всегда. После каждого предъявления стимула испытуемый отвечает, почувствовал он его или нет, и регистрируется процент обнаруженных стимулов каждой интенсивности. На рис. 8.11 показаны примеры полученных таким способом результатов. Точки соответствуют относительным частотам выявления стимулов разных интенсивностей. У большинства испытуемых проходящая через них кривая - психометрическая функция Sобразная. Порогом считается величина стимула, при которой он обнаруживается в 50% случаев. На рис. 8.11 порог не совпадает ни с одним из предъявленных стимулов и определяется путем интерполяции.
S-образная психометрическая функпия часто по форме совпадает с кумулятивной кривой нормального распределения (интегралом гауссова распределения). Если относительные частоты обнаружения случайно предъявляемых стимулов наносятся на график как вероятности отклонения от среднего (величина ζ по правой оси ординат), экспериментальные точки образут примерно прямую линию (рис. 8.1 ,Б). То, что психометрическая функция зачастую описывается S-образной кривой, вызывает определенный теоретический интерес. Значит, в основе флуктуаций в восприятии лежит статистический процесс (см. также следующий раздел).
Статистический анализ порогов. Сенсорная теория принятия решений
В предыдущем разделе мы гипотетически определили абсолютный порог как интенсивность сти-
|
Рис. 8.11. Психометрическая функция, получаемая при измерении пороговой интенсивности посредством случайного предъявления стимулов (метод «постоянных стимулов»). Порог определяется как точка нв кривой, соответствующая обнаружению стимуле в 50% спучвев. А. Зависимость относительной чвстоты успешных попыток рвзличения (ось ординат) от интенсивности стимуляции (ось абсцисс). Б. Зачастую S-образнвя психометрическая функция соответствует интегралу нормального (гауссова) распределения. Если относительную частоту успешных попыток преобразовать в величины Ζ (отклонение от среднего), психометрическая функция принимает вид прямой линии (по [1] с изменениями) |
мула, приводящую к возбуждению сенсорного канала, едва заметно превышающему уровень спонтанной активности его нейронов. Если предположить, что флуктуации этой активности и изменчивость ответов на слабый (постоянный и повторяющийся) стимул описываются кривой нормального распределения, то получается частотный график типа показанного на рнс. 8.12, А. Из него ясно, что наблюдатель, у которого возникает нейронное возбуждение на уровне «а», не может с уверенностью судить, соответствует ли оно спонтанной активности или же воздействию слабого стимула. В случае сильных стимулов такой проблемы нет, поскольку распределение ответов на них не перекрывается со спонтанной активностью.
Проблемы идентификации околопороговых стимулов на фоне шума рассматриваются «сенсорной теорией принятия решений» (теорией обнаружения сигналов) [15, 21]. В соответствии с ней, наблюдатель, пытающийся выделить слабый сигнал из шума, должен принять решение. Оно может, например, состоять в том, что все уровни нейронного возбуждения, превышающие «б», нужно считать стимулом, а все более низкие - спонтанной активностью. Разделительная линия, соответствующая такому решению, называется критерием или отклонением. В примере, иллюстрируемом рис. 8.12,А, решения, исключающие ошибку, невозможны. В случаях, соответствующих равномерно закрашенным (красным цветом) зонам, стимулы не обнаруживаются, а в заштрихованной области справа от критерия спонтанная активность ошибочно воспринимается как ответ на стимул.
Дмо-модель: fornit.ru/38103
Источник:
Дата создания: 04.08.2015
Последнее редактирование: 01.06.2024
Относится к аксиоматике: Системная нейрофизиология.
Другие страницы раздела "Возбуждение и торможение":
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя