
Кортикальные колонки и модули
Системная нейрофизиология (nan)
Нейроны вертикальной колонки получают от проекционных нейронов таламуса афферентные сигналы, характеризующие определенную субмодальность. Несколько вертикальных колонок, связанных с разными субмодальностями, образуют функциональный модуль, способный перерабатывать всю информацию, связанную с различными субмодальностями. Информацию от соседних областей перерабатывают другие функциональные модули, организованные по тому же принципу. Взаимодействующие друг с другом модули образуют распределенные системы коры.
Вес уверенности: Имеет дополнительные подтверждения независимых специалистов
Нервные клетки коры, отвечающие на определенный вид раздражения, образуют кортикальные колонки (рис. 41).
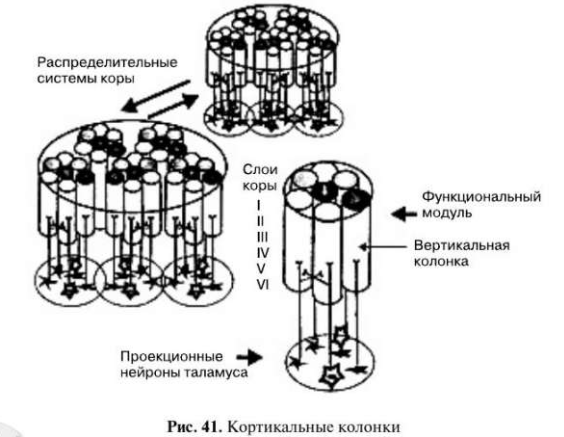
Каждая кортикальная колонка представляет собой ориентированный перпендикулярно поверхности (т.е. вертикально) цилиндр диаметром 300-500 мкм. Нейроны одной колонки получают сходные афферентные сигналы и образуют синапсы преимущественно друг с другом. В колонках проекционной коры имеются две разновидности пирамидных нейронов, которые получили название простых и сложных. Нейроны вертикальной колонки получают от проекционных нейронов таламуса афферентные сигналы, характеризующие определенную субмодальность. Несколько вертикальных колонок, связанных с разными субмодальностями, образуют функциональный модуль, способный перерабатывать всю информацию, связанную с различными субмодальностями. Информацию от соседних областей перерабатывают другие функциональные модули, организованные по тому же принципу. Взаимодействующие друг с другом модули образуют распределенные системы коры.
Простые нейроны, расположенные поблизости от входного IV слоя, выполняют функцию детекторов элементарных признаков раздражителя. Например, простые нейроны зрительной коры сильнее всего реагируют на линейный зрительный стимул в виде полосы или грани между светлой и темной плоскостями, т.е. реагируют на контраст. Друг от друга они отличаются по чувствительности к разным углам наклона линейного стимула. Аксоны нескольких однотипных простых нейронов конвергируют к сложным, или комплексным, нейронам, поэтому комплексные нейроны реагируют на стимул позже простых клеток.
Рецептивное поле сложных нейронов состоит из небольших рецептивных полей простых нейронов, а эффективными раздражителями для комплексных нейронов становится уже не линейный раздражитель определенной ориентации, а характер и направление его движения через рецептивное поле. Так осуществляется переход от детекции простых сигналов к абстрагированию, обобщению. В этом процессе участвуют интернейроны локальной сети кортикальной колонки, а итогом преобразования становится уровень активности выходных нейронов, аксоны которых покидают колонку, чтобы обеспечить ее взаимодействие с другими колонками. Каждая кортикальная колонка представляет собой функциональную единицу коры, или микромодуль.
С помощью горизонтальных связей соседние колонки объединяются в гиперколонку, или модуль. Если микромодули предназначены для распознания элементарных признаков раздражителя или образа, то модуль служит для комплексного объединения элементарных признаков.
Каждый проекционный нейрон таламуса получает информацию от рецепторов одной модальности и возбуждает входные звездчатые нейроны преимущественно одной кортикальной колонки, выбор которой подчинен принципу топографической организации связей сенсорной системы. Соответственно этому, для любой из соседних с этой кортикальных колонок тоже существуют собственные проекционные нейроны таламуса. Аксоны входных звездчатых нейронов оканчиваются в пределах одной кортикальной колонки, тогда как выходные нейроны этой колонки, представленные пирамидными клетками, обеспечивают связи различных участков коры между собой и с другими структурами мозга, в том числе и с таламусом. Таким образом, связь между таламусом и корой является двусторонней, что обеспечивает регуляцию получения информации по принципу обратной связи.
Вторичная сенсорная кора отличается от первичной тем, что большую часть афферентной информации получает не напрямую из таламуса, а из первичной сенсорной коры, а также из других вторичных областей этой же сенсорной системы. Благодаря конвергенции входных сигналов из разных источников здесь происходит по этапная интеграция поступившей от разных рецепторов информации и на основе элементарных свойств раздражителя создается комплексное представление о нем. Участвующие в этом процессе нейроны сильнее всего реагируют именно на определенный комплекс признаков, а не на один из них, как это наблюдается у нейронов первичной коры. Разные функциональные модули вторичной коры могут быть связаны друг с другом ассоциативными волокнами, образованными преимущественно пирамидными нейронами IV—II1 слоев. Если модули находятся на большом расстоянии друг от друга, такие волокна проходят в составе белого вещества. Модули, расположенные в разных полушариях, могут передавать информацию посредством волокон мозолистого тела (межполушарной связи). Волокна пирамидных нейронов обычно группируются в виде пучка, в составе которого аксоны направляются к клеткам-мишеням. В модулях ассоциативной коры отсутствуют клетки, реагирующие на элементарные раздражители, но имеются нейроны, разряжающиеся только в ответ на определенный сложный комплекс стимулов. Например, в нижней височной доле существуют нейроны, которые активируются только при зрительном восприятии человеческого лица.
Ассоциативная кора получает входную информацию от разных сенсорных систем, причем большую часть этой информации поставляют вторичные сенсорные области. При участии подкорковых структур мозга здесь формируются полноценное комплексное восприятие реального окружающего мира и воспоминания о нем. На каждом из иерархических уровней переработки сенсорной информации в коре существуют функционально отличающиеся области, которые специализируются на разных сторонах процесса восприятия.
Источник:
Дата создания: 26.08.2020
Последнее редактирование: 26.08.2020
Относится к аксиоматике: Системная нейрофизиология.
Другие страницы раздела "Нейронные сети":
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя