Ознакомьтесь с Условиями пребывания на сайте Форнит Игнорирование означет безусловное согласие. СОГЛАСЕН
Книги сайта: «Мировоззрение»,
«Познай себя» , «Основы адаптологии» ,
Фантазмы , Лекторий МВАП , «Что такое Я» , «Наука», «Субъективность».
Чтобы оставлять сообщения нужно авторизоваться.
Тема форума: «Эксклюзивные занятия по МВАП: Первые управляющие структуры организмов на основе распознавателя.»
| Сообщений: 135 Просмотров: 9120 | | Вся тема для печати |
1 2 3 4 5 6 7 8 9
nan - админ
- админ Род:  Сообщений: 12298 личная фото-галерея  Оценок: 39 Оценок: 39список всех сообщений clons Сообщение № 46240 показать отдельно Ноябрь 11, 2019, 08:06:29 AM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа:  Благодарность от: Айк, Клон Благодарность от: Айк, Клон | |||||||||
|
Айк
- админ Сообщений: 3768 !!! личная фото-галерея Оценок: 4список всех сообщений clons Сообщение № 46247 показать отдельно Ноябрь 12, 2019, 08:05:39 PM ответ -только после авторизации | |||||||||
Метка админа: | |||||||||
|
nan
- админ Род: Сообщений: 12298 личная фото-галерея Оценок: 39список всех сообщений clons Сообщение № 46248 показать отдельно Ноябрь 12, 2019, 08:17:19 PM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа: Благодарность от: Айк | |||||||||
|
Palarm
- админ Род: Сообщений: 2771 личная фото-галерея Оценок: 6список всех сообщений clons Сообщение № 46257 показать отдельно Ноябрь 14, 2019, 06:22:19 PM ответ -только после авторизации | |||||||||
Метка админа: | |||||||||
|
nan
- админ Род: Сообщений: 12298 личная фото-галерея Оценок: 39список всех сообщений clons Сообщение № 46258 показать отдельно Ноябрь 14, 2019, 07:02:36 PM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа: Благодарность от: Айк | |||||||||
|
nan
- админ Род: Сообщений: 12298 личная фото-галерея Оценок: 39список всех сообщений clons Сообщение № 46260 показать отдельно Ноябрь 16, 2019, 09:26:03 AM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа: Благодарность от: Айк, Клон | |||||||||
|
Клон
- unlimited Род: Сообщений: 1177  личная фото-галерея Оценок: 5список всех сообщений clons Сообщение № 46262 показать отдельно Ноябрь 16, 2019, 11:47:33 PM ответ -только после авторизации | |||||||||
Метка админа: | |||||||||
|
Palarm
- админ Род: Сообщений: 2771 личная фото-галерея Оценок: 6список всех сообщений clons Сообщение № 46263 показать отдельно Ноябрь 17, 2019, 08:25:42 AM ответ -только после авторизации | |||||||||
Метка админа: Благодарность от: nan, Клон, Айк | |||||||||
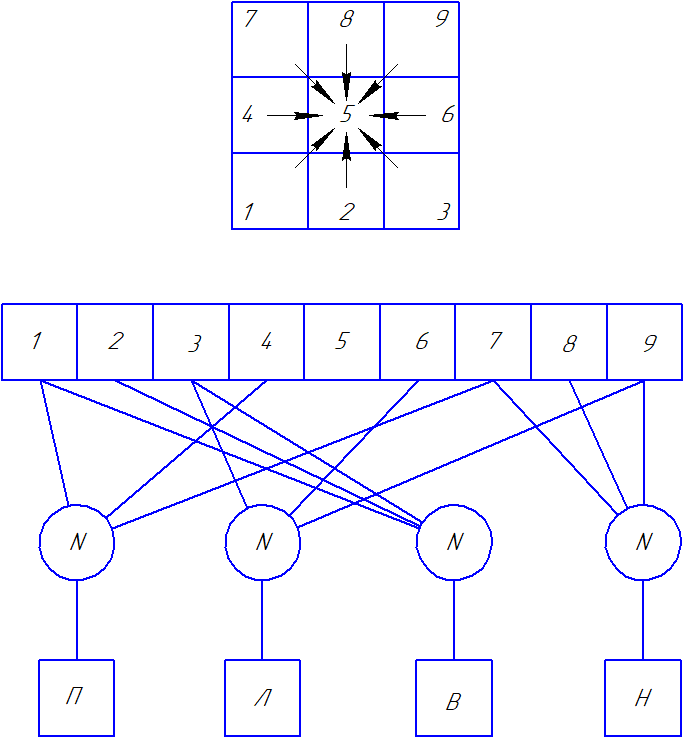
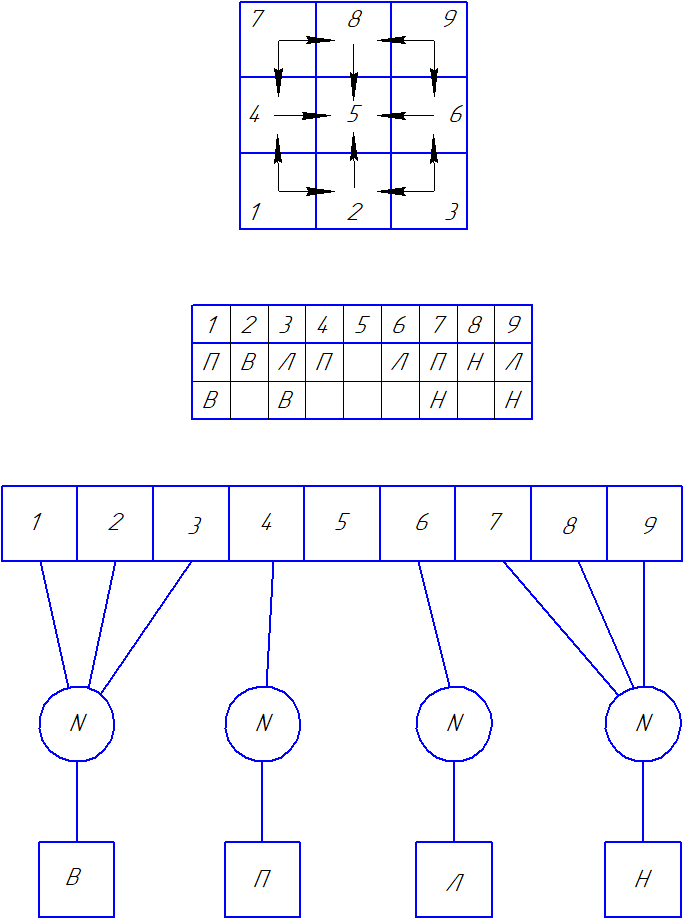
|
nan
- админ Род: Сообщений: 12298 личная фото-галерея Оценок: 39список всех сообщений clons Сообщение № 46264 показать отдельно Ноябрь 17, 2019, 09:00:06 AM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа: | |||||||||
|
Palarm
- админ Род: Сообщений: 2771 личная фото-галерея Оценок: 6список всех сообщений clons История редактирования (2)
Сообщение № 46265 показать отдельно Ноябрь 17, 2019, 09:19:04 AM ответ -только после авторизации | |||||||||
Метка админа: Благодарность от: nan, Клон, Айк | |||||||||
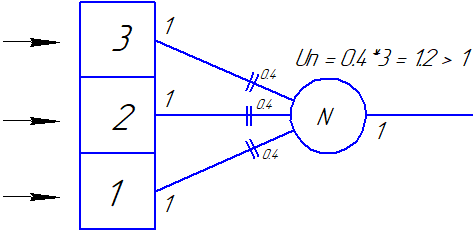
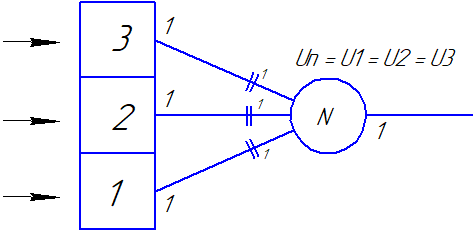
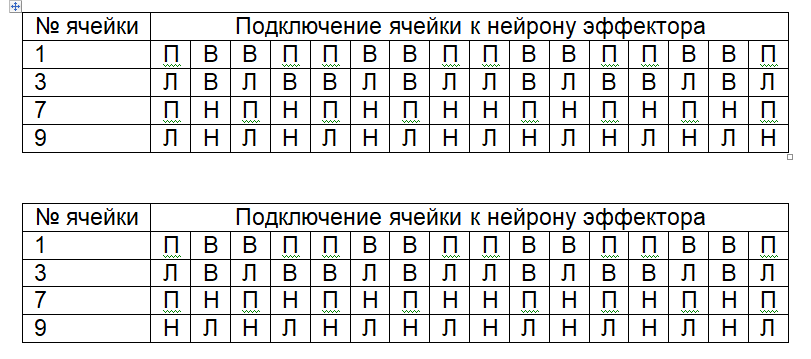
|
nan
- админ Род: Сообщений: 12298 личная фото-галерея Оценок: 39список всех сообщений clons Сообщение № 46266 показать отдельно Ноябрь 17, 2019, 11:59:06 AM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа: | |||||||||
|
Palarm
- админ Род: Сообщений: 2771 личная фото-галерея Оценок: 6список всех сообщений clons Сообщение № 46267 показать отдельно Ноябрь 17, 2019, 01:42:10 PM ответ -только после авторизации | |||||||||
Метка админа: Благодарность от: Айк | |||||||||
|
Palarm
- админ Род: Сообщений: 2771 личная фото-галерея Оценок: 6список всех сообщений clons Сообщение № 46268 показать отдельно Ноябрь 17, 2019, 01:52:53 PM ответ -только после авторизации | |||||||||
Метка админа: | |||||||||
|
Palarm
- админ Род: Сообщений: 2771 личная фото-галерея Оценок: 6список всех сообщений clons Сообщение № 46272 показать отдельно Ноябрь 20, 2019, 01:28:15 PM ответ -только после авторизации | |||||||||
Метка админа: | |||||||||
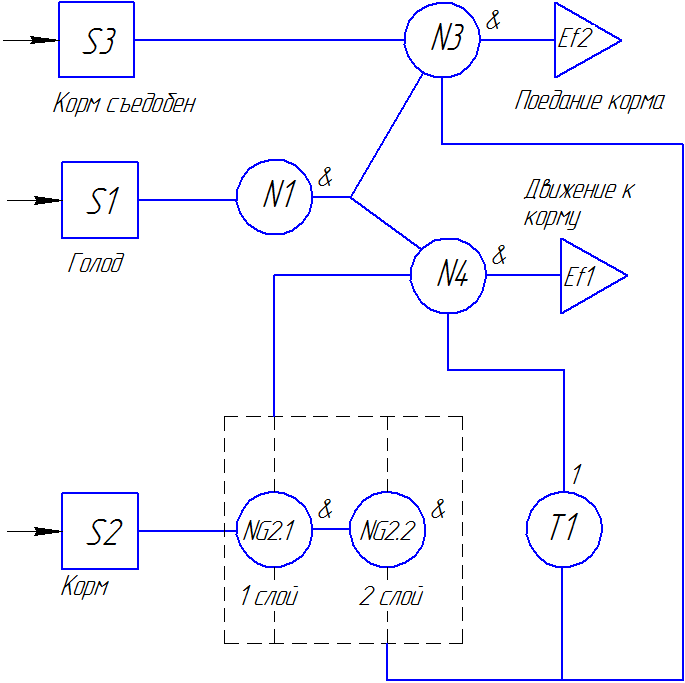
|
nan
- админ Род: Сообщений: 12298 личная фото-галерея Оценок: 39список всех сообщений clons Сообщение № 46273 показать отдельно Ноябрь 20, 2019, 05:58:48 PM ответ -только после авторизации | |||||||||
p.s. Допускаю, что мое утверждение может быть неверно, поэтому прошу показывать, что именно и почему неверно и запрашивать объяснения, если что-то непонятно. Метка админа: | |||||||||
Всего Тем: 1928 Всего Сообщений: 47875 Всего Участников: 5219 Последний зарегистрировавшийся: Geo
Страница статистики форума | Список пользователей | Список анлимитов
Последняя из новостей:
Страница статистики форума | Список пользователей | Список анлимитов
Эгоцентризм – основа субъективности. Когда механизм становится переживанием? Две половинки мозга и одна субъективность: Формирование субъективного опыта в онтогенезе.
Пользователи на форуме:| Обнаружен организм с крупнейшим геномом Новокаледонский вид вилочного папоротника Tmesipteris oblanceolata, произрастающий в Новой Каледонии, имеет геном размером 160,45 гигапары, что более чем в 50 раз превышает размер генома человека. | Тематическая статья: Формирование субъективного опыта в онтогенезе |
Рецензия: Работы А.Иваницкого, комментарии | Топик ТК: Свойства осознанного внимания и циклы |
| Из коллекции изречений: >>показать еще... |