Доказано, что разнообразие растительных сообществ поддерживается расхождением видов по разным нишам
Шалфей Salviacolumbariae. Этот использованный в опытах вид, при отсутствии расхождения видов по нишам, через 20 лет должен был бы стать абсолютным доминантом, на который приходилось бы 99,9% всех растений. Фото с сайта www.laspilitas.com
Путем кропотливых экспериментов с искусственно сформированными сообществами растений-однолетников ученым впервые удалось получить прямые доказательства того, что расхождение разных видов растений по разным экологическим нишам — это реально действующий механизм поддержанию высокого видового разнообразия сообществ.
В последнее время на страницах ведущих научных журналов идут жаркие споры о том, должны ли виды, обитающие в одном месте (и при этом конкурирующие за одни и те же ресурсы), занимать разные экологические ниши. Согласно традиционным воззрениям (принципу конкурентного исключения Гаузе), расхождение видов по разным экологическим нишам — обязательное условие их сосуществования. Однако экологи, изучающие растительные сообщества, не раз обращали внимание на то, что для растений возможности расхождения видов по разным нишам в принципе довольно ограничены. Число совместно произрастающих видов в реальности может во много раз превышать число факторов, лимитирующих рост популяций отдельных видов («измерений ниши»).
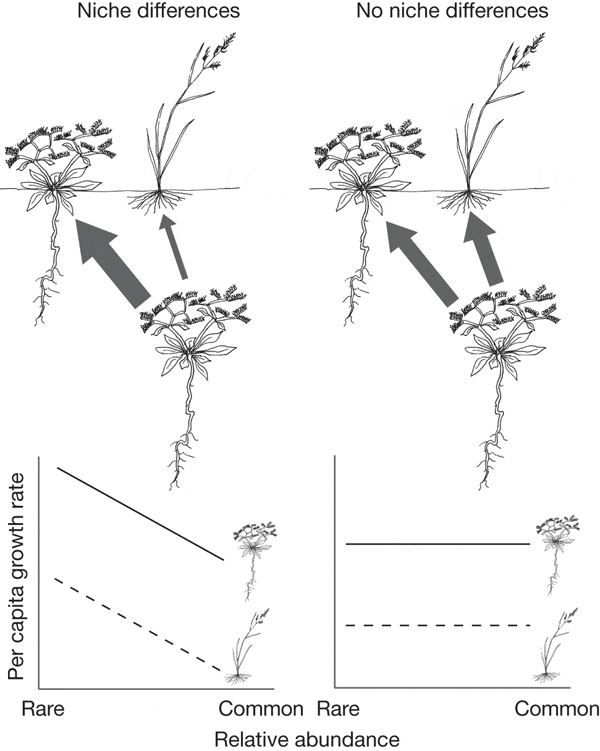
Особенно впечатляет разнообразие деревьев во влажных тропических лесах, где на одном гектаре может произрастать более сотни разных видов, хотя все они конкурируют за одни и те же ресурсы, прежде всего за свет. Неудивительно, что изучение именно таких лесов заставило американского эколога Стивена Хаббела () выдвинуть концепцию нейтрализма, согласно которой разные виды растений могут сосуществовать благодаря не расхождению своих ниш, а наоборот — благодаря их сходству. Если согласно нишевой концепции при возрастании численности популяции какого-либо вида относительно других видов его удельная (в расчете на особь) скорость популяционного роста должна снижаться, то нейтралистская модель предполагает, что эта скорость остается неизменной (см. два нижних графика на рис. 1).
Рис. 1. Схема взаимоотношений двух видов растений, занимающих разные экологические ниши (располагающие корни на разной глубине). Различие в нишах (левый верхний рисунок) проявляется в том, что при увеличении численности популяции одного вида конкурентное воздействие на особей своего вида оказывается сильнее (показано более толстой стрелкой), чем на особей другого вида (тонкая стрелка на том же рисунке). Если же различия в нишах не существенны (правый верхний рисунок), то при росте численности одного из них сила конкурентного воздействия остается одинаковой для особей как своего, так и чужого вида (стрелки одинаковой толщины на правом верхнем рисунке). Графики внизу показывают, как зависит удельная (в расчете на особь) скорость роста популяции растения от его относительной численности (relative abundance) в сообществе. Левый нижний график предполагает расхождение ниш: удельная скорость популяционного роста снижается по мере того как вид из редкого (rare) становится массовым (common). Правый нижний график предполагает отсутствие расхождения видов по нишам: удельная скорость популяционного роста не зависит от относительной численности вида, одинакова как для случая, когда вид редок, так и случая, когда вид обычен. Рис. из обсуждаемой статьи в Nature
Подтвердить гипотезу нейтрализма (как, впрочем, и противоположную ей гипотезу обязательного расхождения видов по нишам) путем прямых экспериментов довольно сложно. Поэтому обычно исследователи ищут косвенные пути проверки. Например, строят математические модели, исходящие из тех или иных предположений об особенностях видов, а затем сравнивают прогнозируемое моделью соотношение численностей разных видов в сообществе с тем, которое реально наблюдается в природе (см: ).
Заповедник Седжвик в Калифорнии, где проводили свои исследования Джонатан Левин и Дженнике Хилрисламберс. Видна экспериментальная площадка, на которой располагались делянки. Фото с сайта protist.biology.washington.edu
Однако недавно два исследователя из Отдела экологии, эволюции и морской биологии университета в Санта-Барбаре (, Santa Barbara, California) Джонатан Левин () и его бывшая аспирантка Дженнике Хилрисламберс () предприняли смелую попытку экспериментальным путем проверить гипотезу, согласно которой высокое видовое разнообразие сообществ поддерживается за счет расхождения видов по разным нишам.
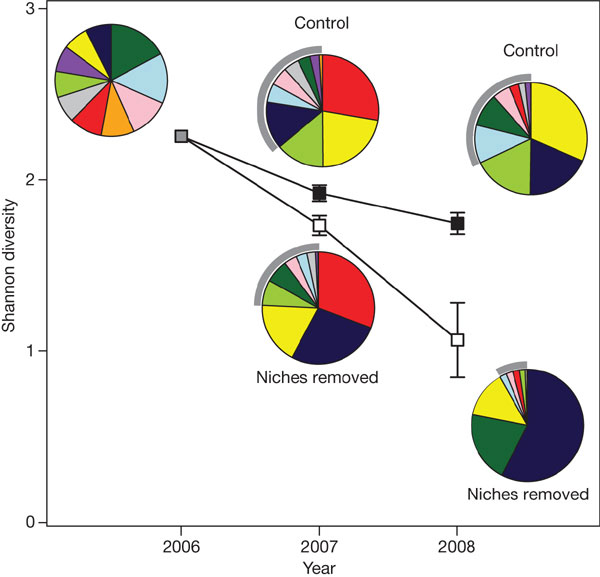
Изменение соотношения видов (по количеству образованных семян) и падение индекса видового разнообразия (Shannon diversity) — показателя, учитывающего как число видов, так и равномерность соотношения численностей всех видов сообщества за два года исследований. Две нижние диаграммы (за 2007-й и 2008 год) — при отсутствии разделения ниш (Niche removed). Две верхние диаграммы за 2007-й и 2008 год — контроль (Control). Диаграмма в верхнем левом углу — исходное соотношение видов, заданное экспериментаторами. Разным цветом показаны разные виды. Серой дугой показано обилие семян 7 наиболее редких видов (из 10 использованных для опыта). Рис. из обсуждаемой статьи в Nature
Navarretia atractyloides. На основании популяционной динамики 2007 года этот вид, при отсутствии разделения по нишам, должен был бы удвоить свою численность на следующий год. Фото с сайта calphotos.berkeley.edu
Объектом их исследований стали искусственно формируемые сообщества из мелких растений-однолетников, развивающихся на так называемые серпентиновых почвах (содержащих труднорастворимые, медленно разрушающиеся силикаты магния, см.: ). Поскольку район исследований — около Санта-Барбары, Калифорния — характеризовался средиземноморским климатом с сухим жарким летом и мягкой влажной зимой, семена растений-однолетников, находящиеся в почве, начинали прорастать в конце осени — начале зимы, а выросшие из них растения сами давали семена весной или в начале лета. Растения эти небольшого размера — на площади 1 м2 их может произрастать около 2,5 тысяч, а разнообразие при этом довольно высокое — на участке 25 × 25 см2 можно насчитать более десятка видов.
Самым трудным в данной работе было свести к возможному минимуму влияние расхождения видов по разным нишам. Авторам пришлось комбинировать эксперименты и математическую модель роста однолетников, причем параметры модели определяли исходя из прямых наблюдений за посевами однолетников в течение двух вегетационных сезонов: 2006–2007 г. и 2007–2008 г. (второй год был более влажным). Всего было отобрано 10 разных видов (представителей разных семейств), обычных для данной местности. Их высевали на специальных делянках, так чтобы суммарная масса всех семян составляла 15 г на 1 м2. Исходно брали равные по весу количества семян всех видов, то есть создавали условия искусственно высокого разнообразия. В вариантах, где предполагалось отсутствие расхождения видов по нишам, всходы пропалывали (снижали плотность популяций), а на следующий год высевали семена разных растений в пропорциях, соответствующих тем, что были получены в предыдущий год.
Micropus californicus. Этот вид наименее конкурентоспособен и, согласно динамике 2007 года, должен был бы в отсутствии разделения ниш сократить свою численность на 98%. Фото с сайта www.coepark.org
Оцененные для всех видов скорости популяционного роста различались в этом случае очень сильно — на порядки величин, что неизбежно должно приводить к быстрому конкурентному исключению одних видов другими. Так, согласно расчетам, шалфей Salviacolumbariae через 20 лет должен стать абсолютным доминантом, на долю которого будет приходиться более 99% от общей численности всех растений. Общее видовое разнообразие сообществ, в которых специально ослабляли эффект разделения ниш, было существенно ниже, чем в контрольных вариантах.
Очень важный результат исследования — экспериментальное подтверждение того, что удельная скорость популяционного роста вида возрастала в тех случаях, когда его относительное обилие снижалось. Таким образом, была реально продемонстрирована ситуация, при которой каждый вид при увеличении его популяционной плотности начинает ограничивать рост собственной популяции в большей степени, чем рост конкурентов.
Источник: Jonathan M. Levine, Janneke HilleRisLambers. // Nature. 2009. V. 461. P. 254–257.
См. также:
1) , «Элементы», 20.02.2007.
2) , «Элементы», 13.11.2008.
3) А. М. Гиляров. От ниш к нейтральности в биологическом сообществе // Природа. 2007. № 11. С. 29–37. (Полный текст — , 700 Кб).
4) А. М. Гиляров. Принцип конкурентного исключения (.)





 http://www.scorcher.ru/xml/news.rss
http://www.scorcher.ru/xml/news.rss