Крушинская Н.Л., Лисицына Т.Ю. ХАРАКТЕРНЫЕ ЧЕРТЫ УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ ДЕЛЬФИНОВ
Печатается по изд.: Крушинская Н.Л., Лисицына Т.Ю. Поведение морских млекопитающих. М.: Наука, 1983. С. 127-166.
Экспериментальные данные, характеризующие условнорефлекторную деятельность
дельфинов немногочисленны и в ряде случаев противоречивы. Первые публикации по
этому вопросу принадлежат; Л. Г. Воронину и его ученикам (Воронин, Козаровицкий,
1969; Стародубцев, 1969; Воронин, 1970; Воронин и др., 1971,
1974).
В работе
Л. Г. Воронина, Л. Б. Козаровицкого (1969) приводится подробный анализ
особенностей высшей нервной деятельности афалины при выработке системы
пищедобывательных условных рефлексов на звуковой раздражитель. Проведенные
исследования показали, что скорость образования положительных пищедобывательных
условных рефлексов у дельфина не отличалась от таковой у других позвоночных
животных. При выработке дифференцировок, переделки сигнального значения
раздражителей, угашении положительных условных рефлексов выявились некоторые
особенности их высшей нервной деятельности. Во-первых, авторами отмечается
сильно выраженная ориентировочно-исследовательская деятельность у афалины,
проявляющаяся в период становления условных реакций и при введении новых агентов
в эксперимент. Во-вторых, дельфину была свойственна неуравновешенность нервных
процессов с преобладанием возбуждения. Этим было обусловлено большое число
межсигнальных реакций, часто носивших характер нападения на рычаг или
агрессивных атак сигнализатора при неподкреплении условного ответа. В-третьих, у
дельфина наряду с неуравновешенностью проявилась инертность нервных процессов,
которая могла принимать черты патологической застойности (дельфин нажимал на
педаль до 400 раз, не получая за это подкрепление). Инертностью нервных
процессов авторы объясняют тот факт, что дельфины с трудом привыкают ко всему
новому и отвыкают от старых привычек. В-четвертых, наряду с явно выраженной
инертностью у дельфина была обнаружена хорошая концентрация их нервных
процессов. Об этом свидетельствует устойчивость выработанных условных рефлексов
в ситуациях, которые могли повлечь срыв высшей нервной деятельности (при
применении сшибки раздражительного и тормозного
процессов).
Л.Г.
Воронин и Л.Б. Козаровицкий отмечают склонность дельфина к развитию
неврозоподобных состояний в период выработки торможения условнорефлекторных
реакций. Так, период стабилизации во время выработки дифференцировки затянулся в
связи с тем, что у афалины произошло нарушение условнорефлекторной деятельности:
она отказывалась от корма, не подходила к установке, «зависая» в углу бассейна.
Эти периоды заторможенности у нее сменялись бурными вспышками двигательной
активности: она на большой скорости плавала по бассейну, била хвостом по воде,
атаковала сигнализатор, рвала зубами паралоновую обшивку на рычаге и т. д. Такое
невротическое состояние у дельфина могло быть достаточно
устойчивым.
К
этому следует добавить, что многие исследователи (McBride, Hebb, 1948; Вуд,
1979; Прайор, 1980) отмечают у дельфинов проявление бурных эмоциональных вспышек
во время проведения экспериментов. Так, афалина совершив ошибку во время опыта
сломала установку и впала в затяжное агрессивное возбуждение (Morgan, цит. по:
Воронин, 1970). Ф. Вуд (1979) сообщает, что однажды во время работы связным в
открытом море известный дельфин по кличке Таффи, не получив положенного
вознаграждения за работу, ударил экспериментатора. В наших экспериментах афалина
Дик после каждого неправильного выбора цели била хвостом плавающего рядом ската.
Приведенные примеры показывают, что, работая с дельфинами, всегда следует
учитывать их эмоциональное состояние, проявление которого может неблагоприятно
отражаться на ходе эксперимента. Исследование динамики образования различных
видов пищедобывательных условных рефлексов на звуковые раздражители было
проведено В. А. Протасовым и др. (1976) на большой группе афалин (15 животных).
Образование условнорефлекторных подходов по сигналу к месту подкрепления у
дельфинов в среднем происходило после 3 — 10 предъявлений, стабилизация
условного рефлекса — после 20 — 60 применений условного сигнала. Образование
инструментального условного рефлекса (нажатие рострумом на рычаг) потребовало
более длительной тренировки для укрепления рефлекса (137 — 168 сочетаний).
Угашение выработанных условных рефлексов наступало после 10 — 30 применений
сигнала без подкрепления. Угашение было нестойким и восстанавливалось в том же
опыте. Проведенные на двух афалинах эксперименты показали, что у них легко и
быстро, как и у большинства высших наземных животных образуются тормозные
дифференцировки, условный тормоз первого и второго порядка, рефлексы второго
порядка (Протасов и др.,
1975).
В
экспериментах (Протасов и др., 1976) была показана возможность образования у
дельфинов динамического стереотипа при частичном подкреплении последовательных
применений условных раздражителей. У афалины был сформирован стереотип подхода
на каждое второе и каждое третье применение условного сигнала. Подход на каждое
четвертое применение сформировать не удалось, так как дельфин перестал покидать
стартовую площадку при применении сигнала. По данным В.А. Протасова и др.
(1975), медленно и трудно у афалин образуются рефлексы с дифференцированием
нескольких положительных условных раздражителей и рефлексы с одновременным
предъявлением положительного и тормозного условного раздражителей, но легко и
быстро, как и у большинства высших наземных животных, у дельфинов образуются
дифференцировки и переделки условнорефлекторных ответов при последовательном
предъявлении тормозных и положительных
раздражителей.
На
основании анализа результатов экспериментов, проведенных на большой группе
афалин, авторы (Протасов, Сергеев, 1975; Протасов и др., 1976) заключают, что по
скорости формирования простых пищедобывательных рефлексов дельфины не отличаются
от других позвоночных животных, что подтверждает выводы, сделанные Л.Г.
Ворониным, Л.Б. Козаровицким
(1969).
Обсуждая
полученные данные, В.А. Протасов и др. (1976) выделяют некоторые особенности
поведения дельфинов при образовании у них условных рефлексов. Они заключают, что
для поведения дельфинов в неволе характерны слабая выраженность
ориентировочно-исследовательской деятельности и склонность к развитию
автоматизмов. Наличие автоматизированных реакций отмечалось авторами в начальный
период обучения, когда животные, слабо реагируя на предъявленные сигналы,
монотонно подходили к манипулятору через определенные интервалы времени или
реагировали только на один из манипуляторов, независимо от сигнального значения
предъявляемого раздражителя; в случае альтернативного выбора у них развивался
«односторонний автоматизм». Предрасположенность дельфинов к автоматизированным
реакциям, как полагают авторы, закреплена экологически и связана со спецификой
охоты на косяки рыбы, при которой обеспечивается «вероятностное
подкрепление».
Как
известно (Купалов и др., 1964; Фирсов, 1972; Крушинский, 1977), односторонний
автоматизм реакций возникает не только у дельфинов, но и у других животных в
период выработки рефлексов, когда животное еще не выделило из комплекса
признаков условного сигнала существенный, по которому происходит обучение.
Односторонний автоматизм также отмечается у разных животных при перенапряжении
нервной системы во время решения трудной задачи с альтернативным выбором
(Крушинский и др., 1972; Крушинский,
1977).
На
основании собственных исследований с дельфинами и данных других авторов мы
полагаем, что автоматизм реакций у дельфинов не следует рассматривать, как
отличительный, признак их поведения, поскольку он возникает функционально как
реакция на трудные
ситуации.
Выводы
В. А. Протасова и др. (1976) о слабовыраженном у дельфинов
ориентировочно-исследовательском рефлексе явно вступают в противоречие с мнением
большинства исследователей, изучавших поведение афалин (McBride, Hebb, 1948;
Tavolga, 1966; Воронин, Козаровицкий, 1969; Caldwell, Caldwell, 1972a, b; Вуд,
1979; и многие
др.).
Мнение о
слабовыраженном у дельфинов ориентировочно-исследовательском рефлексе могло
сложиться у экспериментаторов (Протасов и др., 1976) при проведении опытов в
период адаптации животных к условиям неволи. В этот период, по нашим
наблюдениям, дельфины обычно плавают тесной группой, не реагируют на внешние
раздражители и избегают контактов с незнакомыми объектами. Период адаптации у
дельфинов продолжается разное время (Caldwell, Caldwell, 1972b) и зависит от
того, какие взаимоотношения у них складываются с другими обитателями
бассейна.
Внутригрупповые
отношения могут существенно влиять на их работу при проведении экспериментов и
дрессировке (Pryor,
1975).
Р. Дефран,
К. Прайор (Defran, Pryor, 1981) приводят данные, свидетельствующие, что афалинам
в равной степени свойственны как осторожность (пассивно-оборонительный рефлекс),
так и исследовательская реакция на новые объекты и ситуации. В связи с этим
можно предположить, что отсутствие реакции у дельфинов на новые объекты в
бассейне обусловлено временным подавлением ориентировочно-исследовательского
рефлекса на новую обстановку пассивно-оборонительной
реакцией.
Таким
образом, при анализе результатов исследований, касающихся возможности обучения
дельфинов, следует учитывать индивидуальные особенности психического склада
подопытного животного. К ним относятся уровень эмоциональности, склонность к
установлению контактов с человеком, степень выраженности пассивно-оборонительной
реакции и т. д. В связи с этим следует привести один интересный пример из
практики обучения афалины по кличке Василиса. Этот дельфин отказывался
участвовать в опытах, цель которых состояла в обучении животных различать
звуковые сигналы заданных параметров по стандартной пищедобывательной методике.
Любые попытки в течение длительного времени вовлечь ее в работу оказались
тщетными. Во время опытов с другими дельфинами самка «зависала» в углу бассейна.
Вместе с тем в другой форме эксперимента, построенного на игровой активности,
Василиса проявила большую заинтересованность в работе и продемонстрировала
способность к решению простых логических задач (Крушинский и др., 1972). Следует
отметить, что в дальнейшем афалина Василиса на протяжении многих лет была одной
из лучших цирковых артисток Батумского океанариума (Томилин, 1980). Приведенный
пример наглядно демонстрирует, что дельфины могут проявлять индивидуальную
склонность, определяемую свойствами их психического склада, к тем или иным
экспериментам, которые им предлагает человек в условиях
неволи.
Остановимся
несколько подробнее на разборе экспериментальных данных, полученных при
исследовании формирования цепных условных рефлексов и выработке сложных видов
дифференцировок у дельфинов. В своей повседневной жизни дельфинам приходится
реагировать не только на одиночные раздражители окружающей их среды, но чаще на
целые их комплексы. Дельфины живут в мире звуков, многие из которых являются
коммуникационными сигналами, связанными в определенной последовательности.
Исследованию становления условнорефлекторных связей дельфинов на цепной
акустический раздражитель посвящена работа Л. Б. Козаровицкого и др. (1971).
Эксперименты были проведены на самке афалине по пищедобывательной
условно-рефлекторной
методике.
В
качестве условного сигнала применялся акустический цепной раздражитель,
состоящий из трех тонов: А — 12, Б — 6 и В — 3,2 кГц. Каждый член цепи
действовал изолированно 2 с, а весь цепной раздражитель — 6 с. Реакция дельфина
на рычаг при действии условного сигнала поощрялась
пищей.
Полученные
данные показали, что у афалины легко вырабатывается условный рефлекс на цепной
раздражитель. Этот рефлекс укрепляется так же быстро, как и рефлексы на
одиночные раздражители. В процессе выработки условного рефлекса на акустический
цепной раздражитель у дельфина наблюдалось развитие запаздывательного
торможения. Такого рода закономерность наблюдалась другими авторами при
выработке условного рефлекса на цепи раздражителей у обезьян, собак, кроликов,
голубей и рыб (Воронин, 1947; Ширкова, 1949; Овчинникова, 1955; Бару, 1954; и
др.). Вследствие развития запаздывательного торможения по мере укрепления
рефлекса ответные двигательные реакции у животных отодвигались к концу цепи, т.
е. к моменту пищевого
подкрепления.
Проведенные
Л.Б. Козаровицким и др. (1971) эксперименты показали, что становление условного
рефлекса на цепной раздражитель у дельфинов происходит так же, как и у других
позвоночных
животных.
Особенность
образования цепных пищедобывательных условных рефлексов у афалины исследовалась
Ю.Д. Стародубцевым (1969). Дельфин был обучен последовательно нажимать на 8
педалей, за что получал пищевое вознаграждение. Затем ему был предложен более
сложный вариант задачи: нажимать не на каждую педаль, а только в определенной
последовательности (на 2-ю, 5-ю и на 7-ю педали). При таком режиме подкрепления
у дельфина не удалось сформировать последовательную цепь движений. Дельфин, не
получая пищи при нажатии на любую педаль, продолжал многократно на нее
воздействовать разными способами (сверху, снизу, сбоку), разными частями тела
(рострумом, лбом, грудными и хвостовым плавниками). Сходная задача, предложенная
обезьянам, не вызывала у них затруднений. При формировании цепи движений на
определенную последовательность воздействия на рычаги обезьяны, не получив
подкрепления при воздействии на одну педаль, сразу переходили к другой, а не
продолжали упорно манипулировать с одной, как это делал дельфин. Такая
застойность реакции у дельфинов объясняется Л.Г. Ворониным (1970) инертностью их
нервных
процессов.
Сложный
вид навыка, образованного по принципу цепного рефлекса, удалось выработать у
афалины Дж. Лилли (Lilly, 1967). Цепь этих навыков была следующей: дельфин
нажимал на кнопку — вызов лифта, затем заплывал в расположенную там ванну и,
нажимая рострумом на соответствующую педаль, поднимался к поверхности воды, либо
в здание, откуда его переводили в бассейн. Вместе с тем следует отметить, что не
менее сложные цепи условнорефлекторных движений удается сформировать и у крыс.
Р. Пиррел и Дж. Шерман (Pierrel, Sherman, 1963) обучали крыс производить ряд
последовательных действий: взбираться по спиральной лестнице, опускать разводной
мостик и перебираться по нему; влезать по веревке и, опустившись на подъемнике
на площадку, нажимать на рычаг, после чего они получали пищу. Такие сложные
инструментальные цепи формируются в результате подкрепления отдельных движений,
последовательное выполнение которых приводит к образованию столь сложного
навыка.
Таким
образом, приведенные выше данные показывают, что формирование цепных
двигательных условных реакций у дельфинов происходит так же, как и у других
видов млекопитающих. Обнаруживающаяся некоторая специфика поведенческих реакций
дельфинов в процессе обучения может определяться особенностью протекания их
нервных
процессов.
Общение
дельфинов со своими сородичами осуществляется посредством многообразных и
сложных коммуникационных звуков: зрительных, тактильных и химических сигналов. С
помощью этих сигналов дельфины узнают животных, принадлежащих к своему
сообществу, отдельных индивидуумов в группе, определяют их положение на
иерархической лестнице, выбирают полового партнера, узнают о том, что сородич
нуждается в помощи, предупреждают о своих агрессивных намерениях и т. д. Из
сказанного становится ясно, что дельфины в своей повседневной жизни должны уметь
дифференцировать огромное число сигналов, адресующихся к разным анализаторам. М.
Колдуэлл и Д. Колдуэлл (Caldwell, Caldwell, 1968a, 1972а, с) в исследованиях по
изучению коммуникационных сигналов у дельфинов на большом материале показали,
что каждый дельфин обладает индивидуальным, только ему присущим характером
свиста. Для выяснения вопроса, способны ли дельфины различать индивидуальные
свисты других животных, был проведен специальный эксперимент (Caldwell,
Caldwell, 1972а). Самец афалина должен был нажимать рострумом на рычаг только
при сигналах, издаваемых определенными особями. На магнитофонную ленту были
записаны свисты нескольких дельфинов, издаваемых в разных ситуациях. Эту запись
переписали на другую ленту, включив туда свисты еще нескольких дельфинов.
Подопытный дельфин мог безошибочно выделять свист одного индивидуума из восьми
свистов других животных, записанных на магнитофонную ленту. Причем из множества
сигналов, подаваемых восемью дельфинами, он мог отличать разные типы свистов,
принадлежащие только четырем определенным дельфинам. Этот эксперимент показал,
что дельфины могут узнавать своих сородичей по их индивидуальным свистам,
издаваемым в различных
ситуациях.
Экспериментальное
исследование по выработке дифференцирования акустических сигналов было проведено
(Herman, Arbeit, 1973; Herman, 1980) на самке афалине. Акустические сигналы
предъявлялись последовательно из двух динамиков. Положительный сигнал подавался
в случайном порядке из левого или правого динамика; реакцию дельфина на педаль
подкрепляли только в ответ на действие положительного стимула. Новую пару
сигналов вводили после достижения дельфином определенного критерия при
распознавании предыдущей пары звуков. Из 25 пар звуковых сигналов дельфин
правильно дифференцировал 23 пары. Переделка дифференцирования звуковых сигналов
(изменение положительного стимула) также успешно осуществлялась дельфином. При
этом обнаруживалась явная тенденция к улучшению решения этих задач при их
повторении. Это выражалось в уменьшении числа проб, необходимых для достижения
критерия в каждой новой задаче. По мнению авторов, дельфины усваивают
«концепцию» проводимого с ними эксперимента и могут справляться с задачей с
первых ее применений. Ускорение процесса обучения по мере увеличения
последовательных переделок при различении зрительных стимулов наблюдается и у
других животных: горилл (Rumbaugh, Steinmetz, 1971), крыс и голубей, но не
достигается лишь у рыб (Bitterman, 1965). Переделка дифференцировок
рассматривается М. Биттерманом как один из показателей качественного различия
способности к обучению у разных видов позвоночных. Однако вопрос остается
спорным, поскольку по этому признаку не удается найти качественные различия в
познавательных способностях крыс и таких высокоорганизованных животных, как
обезьяны и
дельфины.
Изучение
способности дельфинов различать геометрические фигуры с помощью сонара
проводились главным образом путем выработки у дельфинов инструментальных
пищедобывательных условных рефлексов. Исследования показали, что дельфины с
помощью отраженных эхосигналов могут различать лоцируемые объекты по их размерам
(Morris et al., 1966; Дубровский, Краснов, 1971), форме (Белькович и Борисов,
1971) и материалу (Evans, Powell, 1967; Дубровский и др., 1970). Мы не ставим
своей задачей изложение данных, характеризующих разрешающие возможности сонара
дельфина. Этому вопросу посвящены специальные обобщающие работы (Яблоков и др.,
1972; Айрапетянц, Константинов, 1970; Белькович, Дубровский, 1976). Данные по
эхолокационному различению дельфинами объектов под водой мы будем приводить
только в целях выяснения их познавательных
возможностей.
Одно
из первых и наиболее обстоятельных исследований способности дельфинов к
дифференцированию зрительных стимулов было проведено В. Келлогом, Ш. Райсом
(Kellogg, Rice, 1966) на самке афалине. Дельфину было предъявлено 31 изображение
разных геометрических фигур, из которых составлялись пары для зрительного их
различения. Дифференцируемые фигуры предъявлялись одновременно в двух соседних
прорезях окон специально сконструированного аппарата. При выборе положительного
стимула дельфин дотрагивался носом до изображения соответствующей фигуры и
получал подкрепление. Зрительные стимулы предъявлялись как под водой, так и над
водой. Перед началом опыта, который проводился под водой, дельфин занимал
стартовую позицию перед пластиковым подвижным экраном, раздвижение которого
служило сигналом начала
опыта.
При
одновременном предъявлении дифференцируемых раздражителей более сильный фактор —
пространственное расположение фигур — обычно маскирует более слабый признак —
форму предметов. Для того чтобы упростить задачу различения предъявляемых
зрительных стимулов, В. Келлог и Ш. Раис провели предварительную серию опытов.
Отрицательный стимул, предъявляемый в паре с положительной фигурой, имел
дополнительный контрастирующий признак — большую освещенность и светлую латунную
рамку вокруг изображения фигуры (положительная фигура была изображена на
сплошном черном фоне). По мере тренировки освещенность целей выравнивалась и
рамку у отрицательной фигуры
убирали.
Результаты
экспериментов показали, что дельфин быстро усвоил задачу, и из 25 пар стимулов,
которые предъявлялись под водой, правильно дифференцировал 21 пару
(84%).
В целях
выяснения возможностей дельфинов к переносу опыта было выбрано 12 пар фигур,
которые дельфин различал в результате обучения; 10 пар других комбинаций тех же
фигур предъявлялись дельфину в качестве новых тестовых задач для различения. В
тестовых парах обе фигуры, только положительную или отрицательную, поворачивали
на 90 или на 180°, а в ряде опытов заменяли на другие (рис.
1).
Как следует
из рис. 28, дельфин выбирал положительную фигуру независимо от угла ее поворота
и сочетания в паре с другой отрицательной фигурой, а также при замене
положительного треугольника на круг. Результаты проведенных экспериментов
демонстрируют способность дельфинов к некоторым формам обобщений, сопоставимым,
по мнению авторов, с таковыми у обезьян.
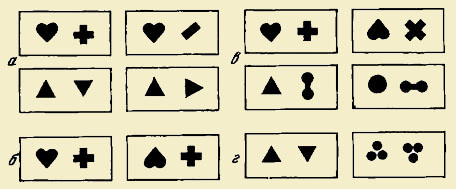
Рис. 1. Примеры изображения фигур, предъявлявшихся дельфину для
исследования его способности к зрительному распознаванию
а — в тестовых парах
изменялась отрицательная фигура; б — изменялась положительная фигура; в —
изменялись обе фигуры; г — задача, не решенная дельфином (по: Kellogg, Rice,
1966)
В той серии опытов, где дельфину фигуры для различения были предложены на
воздухе, он не смог обучиться различать их из-под воды даже после 100
применений. Авторы считают, что это обусловлено искажением изображений фигур
вследствие рефракции. Однако после того как животное было обучено различать
фигуры в воде, оно стало их дифференцировать и на воздухе. Можно думать, что
сложности, возникшие с различением дельфином фигур, предъявляемых на воздухе,
были обусловлены их относительно небольшими размерами (4—5 см2),
близкими к порогу различения. По наблюдениям самих авторов и опытам других
исследователей (Adler, Adler, 1978; Близнюк, 1978; Прайор, 1981), дельфины могут
различать геометрические фигуры больших размеров (30X50 см), предъявленные им на
воздухе, и дифференцировать большое число жестов тренера, которые служат им
сигналами для выполнения определенных
действий.
Исследования
способности дельфинов различать зрительные стимулы под водой были проведены
также Л. Германом и др. (Herman et al., 1969) на одной самке афалине. Различные
зрительные стимулы дву- и трехмерные геометрические фигуры, различающиеся по
форме и размерам; контрастные фигуры — белый круг на черном фоне и черный круг
на белом фоне) одновременно предъявлялись дельфину для различения. Все фигуры
демонстрировались за стеклянным окном в целях исключения возможности
использования дельфином сонара. Как заключают авторы, образование навыка
дифференцирования данных стимулов происходило с трудом и число правильных
ответов не превышало
60%.
Обсуждая
исследования В. Келлога и Ш. Раиса, Л. Герман (Herman, 1980) высказывает
сомнение в том, что в их опытах была надежно исключена возможность
эхолокационного различения предъявляемых стимулов. Так, по его мнению,
особенности рельефа самих латунных пластин, на которые наносился трафаретный
рисунок фигур, могли служить для дельфина источником отраженных эхосигналов.
Приводимые Л. Германом аргументы не кажутся нам достаточно убедительными для
того, чтобы опровергнуть трактовку опытов В. Келлога и Ш. Раиса как обучение
дельфина зрительному различению
фигур.
Несмотря
на некоторые методические погрешности, работа В. Келлога и Ш. Раиса остается
одним из наиболее интересно задуманных и тщательно выполненных исследований
способности дельфинов к сложным видам обучения дифференцированию геометрических
фигур и переносу навыка для выявления отношений между
стимулами.
К
группе сложных дифференцировок относится так называемая методика «выбор по
образцу». В этой задаче животное должно выбрать из нескольких объектов один,
соответствующий сигналу-образцу, на который у него выработан условнорефлекторный
ответ.
В опытах
на обезьянах и дельфинах применяют два варианта этой задачи: выбор по образцу
наличного раздражителя и оставленного во времени раздражителя, т.е. в последнем
случае животное производит выбор по
памяти.
Впервые
эти эксперименты были проведены на шимпанзе Н. Н. Ладыгиной-Котс (1923) в целях
выяснения функций обобщения у обезьян. В этих опытах было выяснено, что для
шимпанзе большее значение имеет признак цвета и меньшее — величины и формы. В
дальнейшем различные варианты «выбора по образцу» применялись многими
исследователями на разных видах обезьян (Маркова, 1967; Фирсов, 1972; и др.) и
продемонстрировали большие возможности этих животных к
обобщению.
Исследование
наличного выбора по тождеству сигналов было проведено в нескольких работах на
афалинах (Антоненко, Близнюк, 1978; Протасов и др., 1976; Adler, Adler,
1982).
В. В.
Антоненко, Я. И. Близнюк (1978) провели подобные исследования на двух афалинах.
Одной предлагалось выбрать из разных по форме, величине и материалу объемных
фигур, находящихся под водой, образец, сходный только по форме. Второму дельфину
предлагалась похожая задача, но фигуры были плоскими и предъявлялись над водой.
При возможности одновременного использования зрительного и эхолокационного
анализаторов у первого дельфина был относительно быстро выработан правильный
ответ при дифференцировке формы предметов. В тех случаях, когда дельфинам
приходилось решать задачу выбора по образцу, пользуясь только зрительным
анализатором (выбор над водой) или только эхолокационным (при очень мутной
воде), выработка такой дифференцировки происходила с большим трудом. Авторы
заключают, что обучение дельфинов сложным дифференцировкам типа «выбора по
образцу» является для них сложной задачей. К такому же выводу пришли В.А.
Протасов и др. (1976), исследуя у двух афалин возможность решения задачи «выбора
по образцу» объемных фигур, предъявляемых дельфинам для локационного различения
под водой. Отмечается невысокий уровень адекватных ответов у дельфинов (69%) при
решении этой
задачи.
Как
следует из этих опытов, существенным облегчением решения задачи выбора по
тождеству предметов для дельфинов была возможность одновременно использовать
зрительный и слуховой
анализаторы.
О
взаимодействии зрительного и слухового анализаторов у дельфинов при различении
целей свидетельствуют специально проведенные (Багдонас и др., 1970) эксперименты
на обыкновенном дельфине при выработке дифференцирования двух геометрических
фигур, различающихся по оптическим и акустическим свойствам и предъявляемых в
разных условиях видимости. Было выяснено, что в ухудшенных условиях для
функционирования эхолокационного анализатора (применение акустически прозрачных
плексигласовых целей) дельфин преимущественно пользуется зрением при различении
фигур. В условиях, затрудняющих зрительное различение (повышенная мутность воды
до 0,5 м), дельфин включал эхолокационный
анализатор.
Приведенные
данные дают основание считать, что при распознавании объектов под водой
зрительный и локационный анализаторы у дельфинов тесно взаимосвязаны и дополняют
друг
друга.
Другим
вариантом задачи выбора по образцу можно рассматривать «смену принципа выбора».
Эта задача была предложена двум афалинам (Adler, Adler, 1982). У дельфинов
вырабатывался условный положительный рефлекс на белый квадрат определенного
размера, предъявляемый над водой, а затем им предлагалось выбрать этот
положительный стимул из четырех других квадратов. В первом варианте опыта они
выбирали его среди четырех белых квадратов разного размера, во втором — из
четырех квадратов такого же размера, но разных оттенков. Оба дельфина с высокой
степенью надежности справились с обоими вариантами этой задачи. Авторы полагают,
что этот эксперимент может демонстрировать достаточно высокий уровень
познавательных способностей дельфинов
афалин.
Исследование
возможности дельфинов решать задачи отсроченного выбора по образцу акустических
сигналов было проведено в большой серии опытов (Herman, Gordon, 1975; Herman,
1975, 1980). Авторами было показано, что дельфины легко справляются с этой
задачей и могут производить выбор по тождеству сигналов с паузой до 8 — 10 с.
Подробное изложение этих экспериментов приводится в разделе «Характеристика
памяти
дельфинов».
Приведенные
выше данные свидетельствуют о том, что дельфины могут проявлять высокий уровень
способности к выработке тонкого дифференцирования зрительных и слуховых сигналов
и при опознавании объектов под водой используют как зрительный, так и
эхолокационный анализаторы, функционирование которых дополняют друг друга при
ориентации животного. Решение дельфинами сложных задач по дифференцированию типа
«выбора по образцу» и вычленение положительной фигуры при пробах на
инвариантность (изменение ее положения, Kellogg, Rice, 1966) показывают, что
афалины, как и обезьяны, способны к обобщениям. Такие формы обобщений Л.А.
Фирсов (1972) склонен рассматривать как «модель элементарной абстракции»,
близкую к интеллектуальной
операции.
При
обучении дельфинов в лаборатории и при дрессировке для цирковых представлений
используют так называемое положительное подкрепление (пищевое, игровое,
тактильное — прикосновение, ласка и т.д.). Методы наказания или отрицательное
подкрепление (удары электрическим током, погружение в холодную воду, действие
сильных звуковых раздражителей и т.д.) обычно при работе с дельфинами не
применяются. Любое неприятное воздействие со стороны человека, в том числе
наказание, надолго нарушает контакт с подопытным животным, что происходит,
по-видимому, в силу их повышенной
эмоциональности.
Вместе
с тем в жизни дельфинам приходится, как и всякому животному, сталкиваться с
воздействием неприятных, вредящих факторов, на основе которых дельфины с детства
обучаются избегать подобные ситуации. Так, для поддержания иерархического
порядка дельфины используют угрожающие звуковые сигналы, позы и прямое
нападение, укусы, удары хвостом, а провинившихся детенышей родители прижимают на
некоторое время ко дну. Трансляция в воду записей голоса косатки оказывала
неизменно паническое действие на стада белух. Этим приемом, как отмечает Ф. Вуд
(1979), пользуются рыбаки для отпугивания стад белух от устья р. Квичак во время
нереста семги.
А.
Марчисон, Р. Пеппер (Murchison, Pepper, 1972) провели экспериментальное
исследование эффективности отпугивающих стимулов на выработку двигательного
условного рефлекса у афалин. В качестве «отрицательного подкрепления» за
неправильное действие применялось сильное звуковое воздействие (акустический
сигнал 50 кГц, создававший давление 63 дБ). По поведению дельфина можно было
установить, что применявшийся звук неприятен для животного. Дельфина помещали в
специальную подводную камеру (4,8X10,8 м), разделенную на две половины невысоким
барьером. При включении акустического сигнала дельфин должен был из стартовой
камеры переплыть в «безопасную зону», которая находилась ближе к неприятному
источнику звука, после чего звук выключался. У дельфина был выработан условный
рефлекс приближаться к подводному источнику неприятного звукового воздействия
для выключения этого звука. В течение первых 100 предъявлений раздражителя,
действовавшего каждый раз в течение 60 с., дельфин переплывал в безопасную зону
в 44% случаев, а при последующих 100 предъявлениях он избегал неприятный
раздражитель в 100% случаев. Этот эксперимент подтверждает возможность обучения
дельфинов в лабораторных условиях по методике выработки оборонительного
рефлекса.
Итак,
исследования формирования условнорефлекторных связей у дельфинов показали, что
по скорости образования положительных пищедобывательных условных рефлексов они
не отличаются от других позвоночных животных. У них вырабатываются положительные
пищедобывательные рефлексы, все виды внутреннего торможения, наблюдается
самовосстановление угашенных реакций, образуются условный рефлекс и условный
тормоз второго порядка. Легко и быстро образуются тормозные дифференцировки,
переделка условных рефлексов. Трудно и медленно вырабатываются, по мнению одних
авторов (Протасов и др., 1976), рефлексы с дифференцированием нескольких
положительных условных раздражителей и рефлексы с одновременным предъявлением
положительного и тормозного условных раздражителей. По данным других авторов
(Kellogg, Rice, 1966), выработка тонких дифференцировок зрительных сигналов с
одновременным предъявлением положительных и тормозных сигналов не вызывает
серьезных затруднений у дельфинов при соответствующей постановке
опытов.
По
основным видам условнорефлекторных связей — положительным условным рефлексам,
некоторым видам дифференцировок, условным рефлексам на цепи раздражителей,
формированию цепных условнорефлекторных движений, переделке сигнального значения
раздражителей выработке активного избегания — дельфины существенно не отличаются
от других высших наземных
животных.
Предъявление
более сложных задач по дифференцированию сигналов типа «выбора по образцу» при
наличном и следовом сопоставлении раздражителей выявило у дельфинов способность
к сложным формам общения. По этим показателям поведения дельфины могут быть
сопоставлены с
обезьянами.
В
период формирования условнорефлекторных связей были выявлены некоторые
особенности их высшей нервной деятельности. Исследователи обращают внимание на
значительную инертность нервных процессов дельфинов, сочетающуюся с их хорошей
концентрацией. Эти особенности обусловливают большую устойчивость временных
связей у дельфинов к действию различного рода дополнительных раздражителей, но,
с другой стороны, затрудняют выработку некоторых временных связей, требующих
быстрого переключения нервных процессов. Одним из отличительных признаков у
афалин многими авторами отмечается значительная неустойчивость нервных процессов
с преобладанием возбуждения, выражающаяся в легком возникновении неврозоподобных
состояний.
Возникающие
у некоторых авторов разногласия при сопоставлении результатов опытов по обучению
дельфинов в большей мере обусловлены тем, что эксперимент обычно проводится на
одной или двух афалинах, обладающих, как известно, индивидуальными особенностями
поведения, которые определяются уровнем эмоциональности, степенью выраженности
ориентировочно-исследовательского рефлекса, пассивно-оборонительной реакции и т.
д. Эти особенности индивидуального проявления высшей нервной деятельности
дельфинов должны учитываться при анализе результатов исследований их
поведения.
В
заключение необходимо отметить, что изучение высшей нервной деятельности
дельфинов требует долгих планомерных исследований с учетом индивидуального
подхода к этим высокоорганизованным животным.
Имитационное поведение
Одним из наиболее интересных и биологически важных видов группового поведения
является подражание, или имитация. Имитационное поведение широко распространено
в животном мире и играет существенную роль в приспособлении отдельных особей и
вида в целом к меняющимся условиям среды. У позвоночных животных подражание
является мощным цементирующим фактором в стадной жизни животных и имеет большое
биологическое значение, поскольку служит основой накопления индивидуального
опыта. Л.А. Орбели (1949) указывал на большое эволюционное значение
имитационного поведения, считая его «главным охранителем
вида».
Исследователями
выделяются две основные категории подражания у животных: стимулирование
видотипичных действий и научение путем подражания или истинное имитационное
поведение (Andrew, 1962; Фабри, 1974). Взаимное стимулирование видотипичных,
врожденных действий широко распространено в животном мире как элемент группового
поведения. В этологической литературе взаимное стимулирование животных
обозначается как аллеломиметрическое поведение. Аллеломиметрическое поведение
проявляется в наиболее простой форме — рефлексе следования. Оно обнаруживается у
стайных рыб, птиц, копытных, хищных млекопитающих, приматов. Одновременный
отдых, собирание пищи, бегство или комфортное поведение животных также являются
примерами аллеломиметрического поведения. Результатом этих действий является
согласованность, унификация и синхронизация поведения членов стада, что
обеспечивает безотлагательное бегство всех животных стада в опасных
ситуациях.
Подлинное
подражание, или «имитационное научение» (Штодин, 1947; Кряжев, 1955), отличается
от аллеломиметрического поведения тем, что животное обучается перенимать те
формы поведения, которые наблюдает у другой особи своего или другого вида.
Имитационное научение проявляется в виде подражания тем формам поведения,
которые не входят в репертуар врожденной видовой деятельности наблюдателя.
Случаи истинной имитации известны для птиц, когда они подражают песне другого
вида или имитируют человеческий голос. Примеры обучения путем наблюдения
известны у рыб (Козаровицкий, 1961), у крыс (del Russo, 1975), кошек (John et
al., 1968), но наивысшими способностями в этом отношении обладают обезьяны
(Воронин, 1947; Кряжев, 1955; Фабри, 1965; Riopelle, Hill, 1973; и
др.).
Формирование
и проявление различных видов подражательного поведения в сообществах животных
находятся в непосредственной связи с иерархической организацией взаимоотношений
членов данной группы. Влияние доминирующих особей в большой степени отражается
на проявлениях подражания в сообществах у обезьян (Menzel, 1971; Воронин, 1947;
Фабри, 1965). Так, опыты Е. Мензеля (Menzel, 1971) с шимпанзе показали, что
группа обезьян, как правило, следует за лидером, которому известно, где
находится тайник с лакомствами, и обычно игнорирует попытки низкорангового
животного увлечь за собой сородичей к известному ему источнику
пищи.
Таким
образом, подражание широко распространено в животном мире и играет существенную
роль в групповых отношениях разных видов. Вместе с тем наивысшего развития
имитационное поведение достигает у приматов и китообразных. Стадная жизнь
большинства видов зубатых китообразных связана с развитием у них разных форм
подражательного поведения. Многие примеры проявления стадного инстинкта или
реакции следования наблюдаются у китообразных в
природе.
Одно из
самых загадочных явлений в жизни китообразных — групповое самовыбрасывание
животных на берег — рассматривается (Norris, 1967; Томилин, 1969) как проявление
реакции следования за гибнущим по той или иной причине членом своего стада. В
отношении причин самовыбрасывания китообразных высказываются различные
предположения: ошибки в работе эхолокационного аппарата, плохо срабатывающего в
условиях мелководья и мутной воды (Dudok van Heel, 1962), неизвестные массовые
смертельные заболевания, психические расстройства (Geraci, 1978; и др.). Однако
ни одна из этих гипотез не получила окончательного подтверждения. Подробное
описание наблюдений за выбрасыванием стада гринд у берегов Флориды приводится в
работе В. Феринга, Р. Велса (Fehring, Welb, 1976). Группа дельфинов, состоящая
из 49 животных, на протяжении четырех дней несколько раз оказывалась на отмели.
Операция по спасению китов крайне затруднялась тем, что отбуксированные в море
животные вновь выбрасывались на берег. Все стадо удалось вернуть только после
того, как далеко в море были отбуксированы два крупных животных, видимо, лидеров
стада. После этого остальные дельфины прекратили попытки, выбрасываться на берег
и стадо объединилось. Однако когда одного из молодых дельфинов спасенного стада
решили взять для океанариума и за веревку, привязанную к его хвосту, начали
подтаскивать к берегу, вновь все стадо последовало за ним. Отдельные животные
стали выбрасываться на берег, разбиваясь о каменные гряды. Даже после того как
молодую гринду оставили, некоторое время дельфины продолжали выбрасываться на
берег. Но и в этот раз удалось спасти оставшихся в живых гринд, отбуксировав их
вместе с лидером далеко в море. Спустя 4 дня часть этого стада (13 животных)
были найдены обсохшими в 275 км от последнего места их обсыхания. Как сообщают
авторы, море в эти дни было спокойным и вода прозрачной, что, видимо, исключает
ошибки в работе эхолокационного аппарата. Среди выбросившихся животных не
наблюдалось паники — они довольно спокойно двигались к берегу и не производили
впечатления больных. Таким образом, во время спасательных операций реакция
следования проявилась у гринд дважды, когда все стадо ушло в море за лидирующими
животными и когда оно последовало за буксируемым к берегу молодым
животным.
Более
сложный вид подражания — обучение путем наблюдений, или истинную имитацию,
исследовали у многих видов зубатых китообразных в неволе: у атлантических афалин
(McBride, Kritzler, 1951; Caldwell et al., 1965), у индийских афалин (Тауler,
Saayman, 1973), у гринды (Kritzler, 1952), у малой косатки (Brown et al., 1966),
у амазонского дельфина (Pilleri et al.,
1980).
Наиболее
обстоятельное изучение различных форм моторного имитационного поведения у афалин
проведено С. Тайлером, Г. Сайманом (Tayler, Saayman, 1973). Две взрослые самки
афалины содержались в одном бассейне со взрослым самцом морского котика
(Arctocephalus pusillus); в этом же бассейне находилось несколько пингвинов,
черепахи и
скаты.
В первые
месяцы их совместной жизни обе самки избегали котика, но постепенно степень их
контактов стала возрастать, в результате чего в поведении афалин появились
многие элементы поведения, присущие котику во время сна, груминга и активного
плаванья. Так, дельфины стали плавать, подгребая плавниками как ластами, а
хвостовой плавник в это время был совершенно
неподвижен.
Как
известно, дельфины те занимаются грумингом. Однако, наблюдая эту форму поведения
у котика, они переняли многие позы и действия, связанные с грумингом. Котики с
помощью передних и задних ласт разглаживают и очищают свой мех, при этом они
принимают разные позы: вращаются вокруг собственной оси у поверхности воды или
принимают вертикальную позу выставив из воды только задние ласты. В это время к
поверхности поднимаются пузырьки воздуха, выдыхаемого котиком. Дельфины,
подражая котику, с помощью грудных плавников научились потирать живот, лежа на
спине у поверхности воды, принимать вертикальную позу, выставив хвостовой
плавник и выпуская при этом пузырьки воздуха из дыхала. Выныривая на
поверхность, они широко раскрывали рот, подражая зевоте котиков, хотя известно,
что зевота не свойственна
дельфинам.
В
первое время своего пребывания в бассейне самки афалин плавали даже во время
ночного отдыха. Котики же спали неподвижно у поверхности воды, лежа на одном
боку, при этом задние ласты выставлены из воды; они имели дугообразную форму.
Иногда они лежали на поверхности на спине, скрестив ласты на брюхе. Спустя
некоторое время дельфины переняли позы котика, которые он принимал во время сна.
Стали замечать, что время от времени афалины неподвижно лежали у поверхности на
боку или на спине, прижав плавники к брюху и выставив хвостовой плавник из воды.
Хотя эти позы были неудобны афалинам, они часто их принимали, находясь рядом со
спящим котиком. Следует отметить, что котик не перенимал поз и движений
дельфинов.
Дельфины
разных видов, содержащихся в одном бассейне, часто перенимают друг у друга
некоторые формы поведения. Так, Д. Браун и др. (Brown et al., 1966) описывают
имитацию прыжков вертящегося дельфина (Stenella lingirostris) афалинами, которые
содержались в том же бассейне. Гринда пыталась балансировать на носу надувную
игрушку, подобно тому как это делали афалины. Но поскольку у гринд отсутствует
вытянутый рострум, она манипулировала игрушкой с помощью
головы.
Некоторые
виды деятельности дельфины, главным образом афалины, могут перенимать у
человека. Так, С. Тайлер, Г. Сайман (Tayler, Saayman, 1973) описывают некоторые
виды орудийной деятельности афалин, которую они наблюдали у людей. Во время
уборки бассейна афалины проявляли большой интерес к этой процедуре. Прибор для
уборки бассейна состоял из полого скребка со шлангом, куда втягивался мусор,
который собирали со дна бассейна с помощью скребка. Однажды аппарат был оставлен
на ночь в бассейне. Им завладела одна самка и долго с ним манипулировала, таская
во рту по бассейну. Затем, подражая действиям человека, она, придерживая шланг
грудными плавниками, взяла в рот совок и стала водить им по дну бассейна. Когда
аппарат убрали, обе самки стали использовать для «уборки» бассейна куски
кафельной плитки (6x8 см), зажимая их зубами. Действуя сообща, они научились с
помощью кафельных плиток собирать в кучу куски морских водорослей
(Enteromorpha), которые они употребляли в пищу. Эти примеры использования орудий
как в форме игры (чистка бассейна), так и для сбора пищевых объектов
свидетельствуют о том, что они могут не только копировать определенные виды
деятельности человека, но и модифицировать их сообразно своим
потребностям.
Имитация
человеческой деятельности свойственна не только афалинам, но и речным дельфинам.
В гл. 2 мы приводили примеры того, как амазонские дельфины, подражая человеку,
«чистили» стенки бассейна и окна с помощью небольших щеточек, которые они
держали во рту (Pilleri et al.,
1980).
Было
проведено (Adler, Adler, 1978) экспериментальное исследование способности афалин
к обучению путем наблюдений на основе использования инструментальной
условнорефлекторной методики. Опыты были проведены на трех афалинах, которые
находились в круглом бассейне, разделенном на четыре отсека крупноячеистой
сеткой. Дельфинам предложили четыре задачи: втягивание свисающего конца веревки
в бассейн; дифференцирование двух квадратов с горизонтальной и вертикальной
исчерченностью; дифференцировку двух квадратов красного и зеленого цвета;
различение цели по ее местоположению (на левой или правой стороне). Все цели
предъявлялись животным над
водой.
С одним из
дельфинов — «демонстратором» проводили эксперимент, а два других находились в
это время в соседних отсеках и могли наблюдать все, что происходит в
экспериментальном отсеке. После закрепления навыка у «демонстратора» одного из
«наблюдателей» переводили в экспериментальный отсек и проводили с ним процедуру
обучения. В опытах те же дельфины были и в роли «демонстраторов» и в роли
«наблюдателей». В последней задаче «наблюдателя» помещали в экспериментальную
камеру спустя четыре дня после проведения опытов с
«демонстратором».
Оценка
результатов опытов показывает, что дельфины могут обучаться сложным задачам
дифференцировать цели путем наблюдений даже с отсрочкой на несколько дней. У
дельфинов-наблюдателей успешное решение задачи происходило уже с первых
предъявлений условных раздражителей. В то же время для обучения
дельфина-демонстратора требовалось большее число тренировок. У «наблюдателей» по
сравнению с «демонстраторами» заметно сокращался латентный период ответов (от 64
до 25 с).
Авторы
полагают, что наблюдения за действиями «демонстратора» уже сами по себе являются
для дельфинов положительным подкреплением всей ситуации и снижают у них
пассивно-оборонительную реакцию перед незнакомыми предметами, которая в сильной
степени развита у этих животных. Л. Г. Воронин (1977) также считает, что
подражательный рефлекс способствует приучению обезьян к экспериментальной
обстановке, сокращая начальный период формирования условнорефлекторной
деятельности.
Приведенные
примеры демонстрируют высокоразвитую способность дельфинов к воспроизведению
сложных моторных действий, но вместе с тем у них в равной степени развита и
«вокальная»
имитация.
Исследования
показали, что дельфины способны имитировать широкий диапазон самых разнообразных
звуков — от булькающих звуков поднимающихся пузырьков воздуха из аквалангов до
отдельных слов и простых фраз, произносимых человеком (Lilly, Miller, 1962;
Caldwell, Caldwell, 1972a; Beach, Papper, 1973; Tayler, Saayman, 1973;
Тарчевская и др., 1978; Richard et al.,
1981).
Изучение
имитации тональных сигналов с частотной модуляцией у афалин было проведено
(Тарчевская и др., 1978) на двух взрослых афалинах. Эталонные сигналы с
частотной модуляцией от 5 до 10 кГц, близкие к естественным свистам дельфинов,
транслировались под водой. Оценка сходства эталонных сигналов с сигналами
дельфинов осуществлялась на экране осциллографа. Было выяснено, что дельфины
имитировали эталонные сигналы свистами, превышающими последние по частоте в 1,2
— 1,8 раз. Иногда дельфины могли повторять эталонный сигнал два-три раза в
пределах одного имитационного. Наблюдалось как непосредственное повторение
только что проигранного сигнала, так и постепенное преобразование собственных
свистов в
имитационные.
Недавно
проведенные исследования способности афалин к имитации искусственных свистовых
сигналов (Richard et al., 1981) показали, что они легко справляются с этой
задачей. Самец афалина, имевший до опытов определенный набор естественных
свистовых сигналов, которые часто и стереотипно повторялись, был обучен
имитировать различные вариации звуковых «моделей», излучаемых генератором. Все
предложенные искусственные звуки дельфин научился точно воспроизводить,
используя свои «вокальные» возможности. Эти звуковые сигналы отличались от его
естественных свистов. При проведении опытов авторы отметили большие возможности
дельфина к обобщению, что позволило ему воспроизводить новые сигналы после 1 — 3
предъявлений. Скорость обучения воспроизведению искусственных звуковых сигналов
свидетельствует, по мнению авторов, о высокоразвитой способности дельфинов
оперировать большим объемом слуховой информации, а способность к «вокальной»
имитации играет существенную роль при установлении коммуникативных связей в их
природных
сообществах.
Широко
известные эксперименты Дж. Лилли (Lilly, 1962), целью которых было изучение
способов установления контактов между человеком и дельфинами, показали, что
дельфины обладают чрезвычайно развитыми «вокальными» подражательными
способностями. Опыты Дж. Лилли подтвердили, что афалина может в какой-то степени
подражать человеческому голосу и после обучения способна воспроизводить слоги,
отдельные слова, простые фразы, восклицания, смех. В своих экспериментах в
качестве подкрепления он применял электрическое раздражение подкорковых структур
мозга, так называемых «центров
удовольствия».
Сравнение
виброграмм с записью голосов дельфинов и человека подтвердило звукоподражание
афалины. Человек произносил слова в пределах колебаний от 200 до 3 000 Гц, а
дельфин воспроизводил их в частотах от 1 000 до 8 000 Гц. Если магнитофонную
ленту с голосом дельфина, имитировавшим слова человека, пропускать с замедленной
в 2 — 4 раза скоростью, то воспроизводится известное сходство с голосом
человека. Дальнейшие эксперименты Дж. Лилли по обучению дельфинов подражанию
человеческой речи, как известно, не подтвердили ожиданий автора о возможности
установления сознательных контактов между дельфином и человеком. Тем не менее
эти опыты продемонстрировали большие возможности дельфинов в сфере имитации
«гомоноидных» звуков, что само по себе можно рассматривать как большую удачу
исследователя.
Интересные
исследования взаимоотношений дельфинов в группе при выработке у них
пищедобывательных условных рефлексов и некоторых форм имитационного поведения
проведены Л. Г. Ворониным и др. (1971, 1974). Группа состояла из трех
черноморских афалин: двух самцов и одной самки. Один самец занимал доминирующее
положение в группе. Было проведено несколько серий экспериментов, где выяснилось
распределение между дельфинами манипуляторов, при воздействии на которые они
сразу получали рыбу. В первой серии опытов дельфины могли нажимать на три
рычага, во второй серии им оставляли только один рычаг, а в третьей — получение
рыбы и манипуляторы были пространственно разделены, т. е. нажав на рычаг,
дельфин должен был плыть за рыбой в другой конец
бассейна.
В
результате проведенных экспериментов было выяснено, что при наличии трех рычагов
внимание дельфинов привлекал в основном тот, который выбирал доминантный самец.
Второй самец предпочитал подбирать рыбу, которая оставалась от работы доминанта.
Насытившись, доминант охотно работал для второго самца, усердно нажимая на
рычаг, и отдавал ему всю рыбу. Самка сначала без всякого предпочтения нажимала
на разные рычаги, но затем чаще стала пристраиваться к самцам и работала обычно
в паре с
доминантой.
При
ситуации, когда оставляли только один рычаг, все животные поочередно на него
нажимали, но чаще других доминантный самец. Несколько раз доминант отгонял
второго самца от манипулятора и преследовал его с агрессивными намерениями. В
том случае, когда работа с манипулятором вознаграждалась в другом месте, чаще
проявлялось деспотическое поведение доминирующего самца. Он отгонял не только
второго самца, но и самку; установилась очередь в работе: сначала к манипулятору
подходил только доминант, затем к добыванию пищи присоединялся второй самец,
потом к работе приступала
самка.
Из
приведенной работы следует, что конкурентные отношения между доминантой и
подчиненным дельфином возникали не по поводу получения рыбы, а из-за доступа к
рычагу, т. е. способу добывания пищи. Таким образом, активный вид деятельности,
приводящий к добыванию пищи, является, по-видимому, прерогативой доминирующего
животного в сообществе дельфинов. Рефлекс подражания у дельфинов данной группы
проявился в том, что наибольшее внимание животных привлекает тот манипулятор,
который выбирал доминирующий самец. Два «свободных» рычага мало интересовали
дельфинов, хотя с их помощью они также могли добывать
пищу.
Обсуждая
результаты проведенных экспериментов, авторы отмечают миролюбие и отсутствие
жестких конкурентных отношений дельфинов в условиях группового эксперимента.
Редко возникающие агрессивные стычки очень быстро сменялись миролюбивыми
взаимоотношениями. Способность доминирующего самца нажимать на рычаг после
собственного насыщения и таким образом «кормить» другого дельфина
рассматривается авторами как проявление сотрудничества в сообществе
дельфинов.
На
основании приведенных в этом разделе данных можно заключить, что у дельфинов в
сильной степени развиты различные формы подражательной деятельности, которые
проявляются в виде реакции следования за лидерами, синхронного плавания, а также
в более сложных формах научения путем подражания. Дельфины имитируют движения,
позы и звуки не только у особей своего вида, но и у животных, далеко отстоящих
от них в таксономическом отношении. Некоторые формы орудийной деятельности,
которые они могут перенимать у человека, можно рассматривать одним из
показателей их развитого интеллекта. Среди млекопитающих по развитию
имитационных способностей дельфины находятся на том же уровне, что и
человекообразные
обезьяны.
Несомненно,
развитая способность к сложным формам подражания играет весьма существенную роль
в социальной жизни дельфинов. Этот вопрос вызывает большой интерес у
исследователей. Л. Герман (Herman, 1980) полагает, что способность дельфинов к
«вокальной» и моторной имитации является той основой, на которой развиваются
различные формы координированной групповой активности: синхронное плавание,
коллективная охота, защитное поведение. Интересную мысль по поводу
функциональной роли имитационного поведения дельфинов в их общественной жизни
высказывают С. Тайлер, Г. Сайман (Tayler, Saayman, 1973). Авторы предполагают,
что имитация половых форм поведения в период ухаживания может играть
существенную роль при выборе полового партнера. Синхронность выполнения
ритуальных поз ухаживания и координация движений во время совместного плавания,
по мнению авторов, может служить стимулом для селективного спаривания и для
упрочения социальных связей в сообществе. Другими словами, в каждом сообществе
могут существовать собственные «традиции» половых взаимоотношений, которые
сохраняются и усиливаются в результате подражания друг
другу.
Развитая
способность к «вокальной» имитации у дельфинов, по мнению Е. Вильсона (Wilson,
1975), может быть причиной большого сходства опознавательных сигналов у
животных, принадлежащих к одному сообществу, что дает им возможность быстро
узнавать своих
сородичей.
Известно,
что у афалин обнаружены (Caldwell, Caldwell, 1972а) устойчивые индивидуальные
«свисты-автографы», представляющие собой определенные контуры частотных
сигналов. Они служат опознавательными сигналами для каждого члена группы. Вместе
с тем можно предположить, что физические характеристики этих
«свистов-автографов» у членов одного сообщества в результате подражания могут
приобрести общие черты, или свойства, отличающие их от подобных индивидуальных
сигналов другого сообщества. По этим сигналам дельфины могут узнавать своих
соплеменников и отличать особей, принадлежащих к другим
сообществам.
Приведенные
выше предположения исследователей о роли имитационных рефлексов в социальной
жизни дельфинов весьма интересны, но требуют экспериментальных
подтверждений.
Память
Изучение памяти является одной из актуальных проблем поведения животных,
поскольку процессы записи, хранения и воспроизведения информации входят
непременным компонентом в любую форму их деятельности — от элементарного
условного рефлекса до образования абстрактных понятий. Исследование особенности
процессов запоминания у животных с высокоорганизованным мозгом представляет
наибольший интерес, поскольку их мозг может хранить и перерабатывать большой
объем информации (Lilly,
1978).
Экспериментальное
изучение памяти у дельфинов стало интенсивно развиваться только в последние годы
(Herman, Bailey, 1970; Herman, Gordon, 1974; Протасов, Сергеев, 1974; Шурепова и
др., 1975; Thompson, 1976; Herman,
1980).
Известны
две фазы памяти: лабильная, или оперативная, которая соответствует удержанию
следа в форме реверберации нервных импульсов (по последним данным, при этом
происходят ультраструктурные изменения нейронов), и стабильная (долговременная),
которой соответствует сохранение следа за счет биохимических изменений в
синапсах (Соколов, 1969,
1981).
Функция
кратковременной памяти состоит в отборе и фиксации новой нужной информации и
оперирование ею в соответствии с требованиями данной ситуации (Baddeley, Hitch,
1974), Оперативная, кратковременная, память играет существенную роль в
формировании адекватных, пластичных и быстрых реакций животных на изменение
окружающей
обстановки.
Основное
внимание исследователей в экспериментах на дельфинах было сосредоточено на
изучении их кратковременной, или оперативной, памяти — той основе, на которой
может развиваться способность к «отвлеченной символизации» предметов и явлений
окружающей их среды (Herman,
1980).
Изучение
кратковременной, оперативной, памяти у афалин проводилось с помощью метода
отсроченной идентификации стандартного стимула — образца при восприятии ими
слуховых и зрительных сигналов (Herman,
1980).
Л. Герман,
Дж. Гордон (Herman, Gordon, 1974) исследовали способность афалин помнить в
течение определенного отрезка времени, какой из двух или нескольких звуковых
сигналов предъявлялся в начале опыта. На один из двух звуковых сигналов у
дельфинов был выработан рефлекс — нажимать на педаль, расположенную у
соответствующего излучателя звука (импульсного или непрерывного). Во время опыта
подавался один из двух сигналов, продолжавшийся 4 с (образец), с некоторой
временной задержкой (от 15 до 120 с) транслировались оба сигнала подряд с
интервалом 0,5 с в случайной последовательности (тестовый сигнал). Нажатие
дельфином педали около излучателя, который передал звук, соответствующий
образцу, считался правильным ответом и поощрялся рыбой (рис.
2).
Дельфину
предлагали разные варианты этой задачи. Так, после предъявления сигнала-образца
через определенный интервал передавали еще один сигнал, который либо
соответствовал «образцу», либо не соответствовал. По тому, какую из педалей
нажимал дельфин, можно было судить, правильно или неправильно он сопоставил
разделенные во времени звуковые сигналы. В наиболее сложном варианте задачи
дельфину предлагалось выбрать стимул-образец из набора разных звуков (Thompson,
Herman, 1977). При проведении этого эксперимента дельфину вначале давали
прослушать серию из восьми новых для него случайно выбранных звуков. Спустя 4 с
транслировали через один из двух боковых излучателей звуковой сигнал. Дельфину
предлагалось решить, совпадает ли этот звук с одним из ранее передаваемой серии
сигналов. Реакция дельфина на педаль, расположенную у излучателя звука, являлась
положительным ответом. Это означало, что «образец» совпадал с одним из сигналов
серии. В том случае, если «образец» не совпадал ни с одним из восьми ранее
передаваемых звуков, дельфин должен был нажать педаль около «молчащего»
излучателя. Дельфин справился с этим сложным экспериментом с высокой степенью
достоверности правильных
ответов.
Зрительная
память у афалин исследовалась (Thompson, 1976) методом сравнения зрительного
образа со стимулами, которые демонстрировались ему спустя определенный
промежуток времени. Все визуальные стимулы предъявлялись дельфину под водой
через застекленное окно размером 46x92 см. Один стимул представлял собой белый
круг на черном фоне (8,3 см в диаметре), второй — черный круг на белом фоне того
же размера. До начала основного эксперимента дельфин был обучен дифференцировать
эти фигуры. В начале опыта ему показывали одну из фигур, а затем сразу или через
определенный интервал времени демонстрировали в окне два стимула. Дельфину
следовало нажать педаль, расположенную у той фигуры, которая демонстрировалась
вначале. Дельфин не мог обучиться сравнивать зрительные стимулы, даже если
«образец» демонстрировался одновременно с тестовыми фигурами, после 100-го
предъявления этой
задачи.
Процедура
обучения выбора по образцу из двух зрительных стимулов значительно облегчилась,
после того как предъявление каждого из зрительных стимулов стали сопровождать
звуками. На первом этапе обучения эти звуки были пространственно разнесены и
проигрывались рядом с изображениями предметов, затем, постепенно сближая
источники звуков, их стали подавать из одного центрального излучателя. После
серии успешных опытов демонстрацию зрительных изображений перестали сопровождать
звуками. Тем не менее дельфин продолжал правильно решать предъявленную задачу
отсроченного выбора зрительных стимулов по образцу. Максимальный интервал во
времени между «образцом» и демонстрацией тестовых изображений не превышал 30 —
35 с.
Л. Герман
(Herman, 1980) объясняет облегчение зрительного распознавания дельфином стимулов
после временного присоединения к ним звуковых сигналов тем, что ассоциативное
обучение осуществлялось в центральном представительстве более высокоразвитой у
них слуховой системы, где зрительные образцы были представлены слуховыми
кодами.
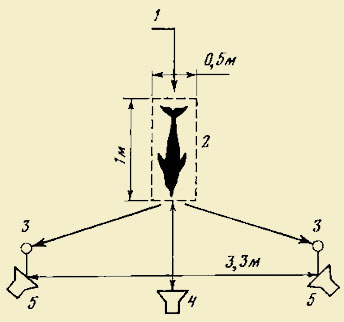
Рис. 2. Схема эксперимента с афалинами «отставленного выбора по
образцу акустических сигналов» 1 — вход в экспериментальный бассейн; 2 —
площадка прослушивания; 3 — боковые педали; 4 — центральный излучатель звука; 5
— боковые излучатели звуков (по: Herman, 1980)
Результаты приведенных выше исследований свидетельствуют о том, что у
дельфинов наиболее развитой оказалась слуховая оперативная память. Это
согласуется с морфологическими данными о более сложном строении слуховых отделов
центральной нервной системы китообразных (Богословская, Солнцева,
1979).
У
дельфинов оперативная слуховая память по сравнению со зрительной обладает
большей емкостью, дольше удерживает новую информацию, отличается пластичностью
при кодировании событий, «считывании» и реализации следовых
процессов.
При
сравнении уровня развития зрительной и слуховой памяти у дельфинов и обезьян
было выяснено (Herman, 1980), что приматы в отличие от дельфинов гораздо лучше
справляются со сложными задачами, где им приходится идентифицировать зрительные
стимулы и плохо обучаются сопоставлять разделенные во времени слуховые сигналы
(Worsham, D'Amato, 1973). Так, опыты, проведенные на капуцинах, в которых
использовался незнакомый для них набор новых предметов, доказали, что обезьяны с
задержкой до 8 мин могли выбирать стимул — образец из набора от 2 до 8 объектов
(D'Amato,
1973).
Сопоставляя
данные обучения дельфинов и обезьян по методу отставленного выбора по образцу,
Л. Герман (1980) приходит к выводу, что зрительная оперативная память у обезьян
столь же хорошо развита, как слуховая у
дельфинов.
Изучение
у собак роли различных рецепторов в продолжительности запоминания комплексного
восприятия местонахождения пищи (Беритов, Айвазашвили, 1967) показали, что
наибольшее значение для сохранения в памяти местонахождения пищи имеет вкусовое
и обонятельное восприятия, меньшее значение имеют зрительное и вестибулярное
восприятия.
Таким
образом, у разных видов животных возможности памяти (восприятие, фиксация и
воспроизведение) определяются уровнем развития их сенсорных систем,
избирательностью выбора и считывания информации со следами предыдущих событий, а
также особенностью эффекторных систем в воспроизведении сигнала (след памяти
может быть воспроизведен лишь в пределах, ограниченных возможностями
исполнительных
органов).
На
основании приведенных выше данных можно заключить, что дельфины успешно хранят и
перерабатывают большой объем, информации, что дает возможность сравнивать их с
приматами. При этом наиболее быстрое, лабильное воспроизведение следовых
процессов и сопоставление их с текущими событиями происходит в пределах слуховой
системы.
Лабораторные
методы исследования, к сожалению, пока не позволяют нам составить более полного
представления о возможностях использования дельфинами ресурсов своей памяти в
естественной жизни. Можно согласиться с мнением Дж. Лилли (Lilly, 1978), что
развитие мозга дельфина обусловлено большим объемом информации, которую он
воспринимает и обрабатывает. В настоящее время можно говорить о трех возможных
источниках необычайно большой по объему информации, которыми могут располагать
китообразные.
Одним
из возможных источников большого объема информации может явиться восприятие
акустических образов окружающих предметов. Это предположение было впервые
высказано Дж. Лилли еще в 1963 г. Предполагается, что мозг дельфинов также
оперирует акустическими аналогами, как мозг человека — зрительными. Сложнейшие
звуковые сигналы дельфинов и других китообразных, которые никак не поддаются
расшифровке, могут содержать не слова и фразы, как у людей, а некое подобие
голограмм. Это также могло бы привести к многократному увеличению передаваемой и
обрабатываемой
информации.
Активная
эхолокация, как полагают В. А. Протасов, Б. Ф. Сергеев (1974), требует развитой
оперативной памяти, так как для получения исчерпывающей информации о подводных
объектах необходимо иметь возможность сравнивать зондирующую посылку с
эхосигналом, возникшим при отражении сигнала от лоцируемых тел. Сложная
социальная организация сообществ у китообразных, поддерживающаяся в первую
очередь с помощью звуковых сигналов при идентификации членов сообщества,
установления контактов внутри группы, поиска полового партнера и т. д., также
связана с восприятием и долговременным хранением большого объема слуховой
информации.
Другим
источником информации можно считать хеморецепцию. С того времени, как Л. И.
Суховской (Suchovskaya, 1972) было доказано наличие вкусовых луковиц на языке
дельфинов, а работами В. Е. Соколова, В. Д. Кузнецова (1971), В. Д. Кузнецова
(1978) была экспериментально подтверждена возможность восприятия малых
концентраций некоторых веществ в воде, предположение о существовании у зубатых
китообразных хеморецепции (Яблоков, 1957, 1961) превратилось в установленный
факт. Учитывая, что запахи в водной среде должны сохраняться много дольше, чем в
воздухе (Яблоков и др., 1972), роль хеморецепции у китообразных по объему
улавливаемой информации может быть сравнима, по мнению автора, с ролью
зрительного анализатора у животных, обитающих на
суше.
Третий
источник информации следует из особенностей социальной организации их
группировок. В каждом стаде дельфинов, видимо, могут находиться одновременно
особи пяти-шести поколений. Этот факт становится очевидным, если учесть, что
продолжительность жизни у большинства видов дельфинов составляет около 25 лет, а
половая зрелость наступает в 2 — 5 лет. Подобное соотношение одновременно
живущих поколений можно встретить только у высших приматов (продолжительность
жизни шимпанзе около 60 лет, а половозрелость наступает к 6 — 10 годам) и
слонов. Указанная особенность состава социальных группировок у китообразных и
высших приматов может в немалой степени способствовать передаче «традиций» и
накоплению большого объема информации от поколения к поколению.
Некоторые подходы к изучению способов общения у дельфинов
Энтузиазм Дж. Лилли, направленный на изучение дельфинов, и его попытки найти
способы общения человека и дельфина (Lilly, 1961 — 1978) побудили многих ученых
заняться этой проблемой. Появились серии работ, в которых исследовались
механизмы звукового общения дельфинов между собой, выяснялось, могут ли дельфины
обмениваться осмысленной информацией, существует ли у них язык символов (Dreher,
Evans, 1964; Lang, Smith, 1965; Busnel, 1966; Bastian, 1967; Batteau, Markey,
1968; Caldwell, Caldwell, 1972 c; Herman, Arbeit, 1973; Белькович, Казнадзей и
др. 1978; Марков, 1978; Herman, 1980; Reiss et al.,
1981).
Одну из
самых интересных работ в этой области выполнил Дж. Бастиан (Bastian, 1967).
Применив методику оперантного обучения, Дж. Бастиан хотел выяснить, может ли
один дельфин, наблюдая за условными оптическими сигналами, сообщить с помощью
акустических сигналов другому дельфину, визуально от него изолированному, какой
из двух манипуляторов следует нажать, чтобы оба получили пищевое поощрение. В
опыте участвовали две афалины — самка и самец, которые были обучены нажимать на
левый или правый рычаг в зависимости от того, какой условный сигнал включался
(мигающий свет или
непрерывный).
В
первой серии опытов дельфинов обучали в разных отсеках бассейна, разделенных
сетью, сквозь которую дельфины могли видеть и друг друга, и сигнальные лампы,
находящиеся в отсеке самки. В отсеке самца над рычагами помещалась лишь одна
лампа, включение которой служило сигналом начала эксперимента. Когда у обоих
дельфинов был выработан условный рефлекс, их разделили перегородкой, через
которую они не видели друг друга, но акустическая связь между ними сохранялась.
Самец мог знать о начале эксперимента по включению лампы в его отсеке, а
условные сигналы (указывающие левый или правый рычаг следует нажимать)
включались только в отсеке самки. Несмотря на то что самец не видел сигнальных
ламп, он вскоре стал правильно (в 90% случаев) нажимать рычаги в соответствии с
тем сигналом, который подавался самке. Изменение сигнального значения условных
раздражителей, т. е. результат «обратного обучения» показал, что и в этом случае
самец стал правильно нажимать на педаль. Только после введения звуко- и
светонепроницаемой перегородки самец стал случайно нажимать на педали.
Результаты столь удачно начатого эксперимента, естественно, привели
исследователей к мысли, что самка с помощью акустических сигналов сообщала
самцу, какой из рычагов, левый или правый, следует нажать, чтобы получить
вознаграждение.
Тщательный
анализ магнитофонной записи звуков самки показал, что самец действительно
ориентировался на звуки, издаваемые самкой при выборе рычагов. Однако
доказательств тому, что самка понимает связь между своими сигналами и поведением
самца, не было получено. После того как самца перевели из отсека в бассейн и
убрали ширму, самка продолжала издавать те же эхолокационные щелчки, на которые
ориентировался самец при выборе манипулятора. Дж. Бастиан полагает, что у самки
в начале опытов случайно или по какой-то неясной причине выработался рефлекс на
включение непрерывного света издавать серию щелчков. У самца также выработался
условный рефлекс на серию щелчков, издаваемых самкой, нажимать на правый рычаг в
то время, когда он мог видеть самку и сигналы лампы. Автор полагает, что в
основе этого сложного согласованного действия лежит самодрессировка самки
издавать разные звуковые сигналы в ответ на включение различных световых
сигналов, а также самодрессировка самца выбирать рычаг в соответствии с
услышанным звуковым сигналом
самки.
Опыты Дж.
Бастиана не дали ответа на вопрос о существовании у дельфинов языкового
смыслового общения, но выясненное в эксперименте умение дельфинов справиться со
сложной задачей выработки тонкой диффереицировки условных сигналов при
совместной работе может свидетельствовать о высоком уровне развития их высшей
нервной деятельности. Данные этих опытов показывают, насколько сильно развит у
дельфинов общественный инстинкт. Звуки, производимые партнером, видимо даже не
относящиеся к коммуникационным, являются достаточно сильными раздражителями, на
основании которых могут формироваться рефлексы второго порядка у членов
сообщества.
Непосредственное
изучение акустических сигналов, которыми обменивались два дельфина вне
визуального контакта, было проведено Т. Лангом, Дж. Смитом (Lang, Smith, 1965).
Они поместили двух дельфинов афалин в разные бассейны и установили между
животными двустороннюю акустическую связь с помощью гидрофонов и подводных
динамиков. Время опыта (30 с) было разбито на 16 интервалов длительностью от 80
до 150 с каждый. В период интервалов с четными номерами между бассейнами
устанавливалась акустическая связь, во время интервалов с нечетными номерами
такая связь отсутствовала. Типы издаваемых свистов были разделены на 6 групп (А
— Е).
Записи
показали, что в периоды отсутствия акустической связи голосовая активность
дельфинов резко падала по сравнению с периодами включения связи. В начале опытов
при отсутствии связи регистрировали свисты трех типов — А, Б, Г. В период
включения связи также присутствовали свисты А, Б, Г и добавлялся еще один тип —
Е. По мере увеличения числа опытов менялся характер звуков в периоды включения
связи. Присутствовали все 6 типов свистов и наблюдалась определенная
последовательность типов свистов: Б — Г — Б — Г. При этом все свисты Б издавала
самка, а свисты Г — самец. Спустя 4 месяца запись звуков, издаваемых самкой,
была несколько раз проиграна самцу. На свист самки Б самец отвечал свистом типа
Г и замолкал каждый раз на том месте, где воспроизводился свист самки Е. На этом
основании авторы сделали вывод, что свисты типа Б и Г — это зов другого
дельфина; свист А — приглашение вступить в голосовое общение. Более сложный
свист Е, по мнению авторов, может нести определенную смысловую нагрузку, так как
этот звук присутствовал только во время двусторонней связи и не воспроизводился
дельфинами при механическом одностороннем
воспроизведении.
Основной
вывод, который можно сделать из проведенных Т. Лангом и Дж. Смитом экспериментов
с двусторонней «телефонной» связью двух дельфинов: они активно обмениваются
звуковыми сигналами, которые, видимо, несут разные функциональные нагрузки. Но
могут ли дельфины передавать с их помощью какую-либо смысловую информацию на
основании этих опытов, сказать нельзя. Итак, ни эксперименты Дж. Бастиана, ни
опыты Т. Ланга и Дж. Смита не выяснили возможности языкового общения у
дельфинов. Главной неразрешенной проблемой в экспериментах Дж. Бастиана остается
возможность влияния дельфинов друг на друга с помощью эхолокационных звуков. До
тех пор, пока не будет устранен этот фактор, трактовка опытов не может быть
однозначной. Е. Н. Панов (1980) высказывает интересную мысль о том, что
объединение обеих методик могло бы внести некоторую ясность в понимание
результатов опытов Дж.
Бастиана.
Было
проведено немало специальных акустических исследований, связанных с попытками
расшифровок языка дельфинов (Dreher, Evans, 1964; Busnel, 1966; Caldwell,
Caldwell, 1968a, 1972c; Белькович, Казнадзей, и др., 1978). Одним из интересных
исследований явились работы В. И. Маркова (Марков, 1978; Марков, Островская,
1978). Авторы выделили в «словаре» афалины 7 исходных сигналов, из которых
формируется 31 элемент более сложного состава. Последние комбинируются друг с
другом, образуя большое количество различных сигналов. В работах В. И. Маркова
продемонстрировано наличие (высокого уровня синтаксической организованности
звуковых сигналов черноморской афалины. У дельфинов было обнаружено до 6 уровней
комбинирования первичных сигналов. Обнаруженная организация звуковых сигналов
подтверждает, что звуковое общение между дельфинами может нести сложную
информацию.
Возможности
установления контактов с дельфинами с помощью свистовых сигналов, передаваемых
человеком, впервые были осуществлены в опытах Д. Батто. Эксперименты Д. Батто,
П. Маркей (Batteau, Markey, 1968), предпринятые с целью выяснения «языковых»
способностей дельфинов, выявили у них удивительные возможности выполнять точные
команды на различные свистовые сигналы. Д. Батто сконструировал электронный
прибор, преобразующий звуки человеческой речи в свистовые, подобные тем, что
издают дельфины. Запись велась постоянно, и по графикам можно было визуально
сравнивать между собой свисты дельфинов и копирующего устройства. Опыты
проводились на двух афалинах. Подавались определенные команды типа: «вперед»,
«направо», «налево», «выйди из обруча», «ударь ластом по мячу» и т. д. За время
работы оба животных научились правильно исполнять 15 таких команд: толкать мяч,
проплывать сквозь обруч, поднимать хвост, издавать эхолокационный щелчок и т. д.
Эти эксперименты показывают, что поведением дельфинов можно управлять с помощью
свистов. Хотя животные воспринимают услышанное не как речь, а просто как
условный звуковой сигнал, они точно выполняли требуемые
действия.
Второй
эксперимент, проведенный с этими же дельфинами, ставил цель выяснить, может ли
дельфин правильно выполнять команду, которой его специально не учили, но он
наблюдал процедуру тренировки со вторым дельфином. На команду «издай
эхолокационный щелчок» не обученный этому ранее дельфин сразу же правильно
отреагировал, издав щелчок. Когда этой же самке подали команду «подними хвост»,
она им шлепнула по воде. Этот опыт еще раз подтверждает удивительно развитые
имитационные способности у дельфинов, сформировавшиеся в результате их стадного
образа
жизни.
Группой
сотрудников Гавайского университета (Herman, Arbeit, 1973; Herman, Gordon, 1974;
Herman, 1980) на трех афалинах были проведены исследования, в которых выясняли
уровень усваивания животными простых языковых звуковых стимулов. Эксперименты
были разработаны только для выяснения возможности восприятия различных
комбинаций звуковых символов, а не для их воспроизведения. Отправной точкой
постановки данных экспериментов было допущено, что у человека в онтогенезе
раньше формируются связи, обеспечивающие понимание языка, чем его
воспроизведение.
Для
дельфинов была разработана кодовая слуховая система, в которой окружающие
дельфинов в бассейне предметы и действия, имеющие к ним отношение, были
обозначены определенными, произвольно выбранными звуковыми сигналами. Эти
сигналы производились с помощью электронного генератора и транслировались в
воду.
В первой
серии опытов дельфина по кличке Ки обучали при демонстрации разных объектов в
бассейне (мяча, спасательного круга и пластикового цилиндра) выбирать один из
них при действии соответствующего этому предмету звукового сигнала. После
закрепления навыка дельфина начали тренировать на выполнение парных звуковых
команд из двух слов, обозначающих объект — действие, например: «коснись мяча»,
«возьми мяч», «принеси мяч». Дельфин в течение 4 месяцев обучился выполнять 9
комбинаций трех действий с тремя предметами. В конце опытов дельфина обучили по
соответствующему звуковому сигналу приносить объекты только определенному
экспериментатору.
В
следующей серии экспериментов участвовали два дельфина. Их тренировали в одном
бассейне. Имя каждого дельфина обозначалось звуковым сигналом. Опыт проходил
следующим образом. Одного из дельфинов по имени вызывали на стартовую площадку.
Услышав «свой» сигнал, он нажимал на педаль, расположенную на его площадке.
Далее следовала команда-инструкция, состоящая из двух слов: существительное —
глагол, сформированная по той же схеме, что и у Ки. Если инструкция выполнялась
дельфином правильно, подавался звук, обозначающий «да» (правильный ответ), когда
дельфин ошибался, включали сигнал
«нет».
В этой
серии экспериментов одного дельфина тренировали на выполнение звуковых команд,
некоторые команды другому дельфину подавались с помощью
жестов.
Словарный
запас дельфинов был доведен до 25 слов (существительных и глаголов). Оба
дельфина оказались способны различать 44 парных комбинаций команд, составленных
из этого словарного
запаса.
После
усвоения парных команд дельфинам стали предъявлять команды, состоящие из трех
слов, например «правый мяч выброси». Дельфин по этой команде выбирал мячик,
расположенный от него справа, и выбрасывал из воды. Трехсложная команда типа
«мяч принеси обруч» обозначала, что дельфин должен поместить мячик рядом с
обручем. По команде «обруч принеси мячик» дельфин должен был отнести обруч в то
место, где находился мяч. Такие сложные команды правильно выполнялись в среднем
в 50% случаев. Это значительно превышает вероятность случайного выбора, так как
число возможных ответов было очень велико. Правильные ответы на 600 — 700 команд
из двух слов составили 80%. Число правильных ответов по одной команде падало с
увеличением числа объектов (6 — 11), из которых производился
выбор.
Проведенные
исследования показали, что дельфины способны в лабораторных условиях к случайно
выбранной элементарной символизации окружающих их явлений и предметов. Однако,
как полагает автор (Herman, 1980), эти эксперименты не означают, что в природе у
дельфинов существует подобная символизация окружающей их
среды.
Недавно
опубликовано сообщение (Reiss et al., 1981) о начале исследований возможности
дельфинов с помощью сложных двигательных навыков оповещать экспериментатора о
событиях окружающей их обстановки в океанариуме. Три афалины были обучены
инициативно использовать кодовые сигналы для получения игрушек, добывания пищи,
установления тактильных контактов с человеком. Во время обучения использовали не
только пищевое поощрение, но и игровое, применяли также некоторые формы
отрицательных подкреплений их нежелательных действий. Первые этапы обучения
прошли
успешно.
Можно
выделить основные направления исследований способов общения у дельфинов. Один
путь — это изучение структуры самих звуковых сигналов, выделение элементарных
исходных единиц — «фонем», анализ последовательности чередования разных типов
сигналов и т. д. Другой подход направлен на выяснение вопроса, могут ли дельфины
обмениваться содержательными сообщениями о характере и расположении окружающих
их предметов, сигнализирующих о возможности получения пищи и др. И третий подход
— это попытки установления контактов между человеком и дельфином с помощью
сигналов-символов, обозначающих предметы и явления окружающей их среды; сюда же
входит обучение дельфинов человеческой речи. Третий подход наиболее широко
используется при изучении «языкового поведения» у обезьян и дельфинов. Попытки
обучения шимпанзе членораздельной человеческой речи (Hayes, Hayes, 1952;
Lieberman, 1968) не привели к желаемым успехам. Исследователи пришли к
заключению, что, несмотря на удивительно развитую способность к подражанию,
голосовой аппарат шимпанзе не приспособлен к воспроизведению речи человека. При
попытках обучения дельфинов человеческой речи (Lilly, 1962) также не были
достигнуты большие успехи, чем при обучении
обезьян.
Опыты Д.
Батто, П. Маркей (Batteau, Markey, 1968), Л. Германа (Herman, 1980) являются
одним из подходов к изучению у дельфинов способности к символизации внешнего
мира. Этот путь исследований, как известно, привел к успешным результатам при
обучении шимпанзе азбуке жестов, используемых глухонемыми в Америке (Gardner,
Gardner, 1972). Исследования Б. и Р. Гарднеров показали, что шимпанзе, обученные
азбуке жестов, могут использовать знаки-символы в качестве средства общения с
экспериментаторами и другими шимпанзе. Шимпанзе по кличке Уошо, выучив несколько
жестов в конкретной обстановке, могла вследствие генерализации употреблять эти
жесты в другой ситуации. Так, жест «открыть», выученный по отношению к ящику с
игрушками, шимпанзе самопроизвольно применяла по отношению к двери и
водопроводному крану. Таким образом, использование шимпанзе их словарного запаса
представляет творческий процесс и не является результатом простого заучивания
слов по методу условного
рефлекса.
Замечательные
исследования супругов Б. и Р. Гарднеров обнаружили у животных способность к
символизации окружающего мира — тех качеств, которые лежат в основе языка
человека. Однако это открытие еще не подтверждает представления о том, что у
шимпанзе реально существует язык, подобный человеческому. Е. Н. Панов (1981)
подчеркивает принципиальное отличие человеческого языка от языка животных: если
язык человека является средством познания мира, то язык животного — лишь
средство
коммуникаций.
Исследования
Л. Германа (Herman, 1980), направленные на изучение уровня способности афалин к
символизации окружающих предметов и явлений с помощью звуковых символов,
находятся на первом этапе. Выяснилась лишь возможность обучения дельфинов
реагировать определенными действиями на различные комбинации звуковых символов.
Цель дальнейших экспериментов, как сообщает Л. Герман, установить двустороннюю
связь человека с дельфином, используя звуковые
символы.
Таким
образом, в настоящее время вопрос о возможности установления двусторонней связи
с дельфинами посредством языковых символов на том же уровне, что и с шимпанзе,
остается открытым. Можно надеяться, что в ближайшее время будет получен ответ на
этот вопрос, уже много лет волнующий
исследователей.
Другим
плодотворным направлением в исследовании коммуникационных возможностей
китообразных является изучение биологического значения сигналов при
взаимодействии животных в естественных группировках. Такие работы в настоящее
время широко развертываются в отношении изучения звуковых сигналов как усатых,
так и зубатых
китообразных.
Возможности
общения с помощью языковых символов у животных тесным образом связаны с уровнем
развития их интеллекта, или рассудочной деятельности. Поэтому в следующем
разделе мы остановимся на некоторых экспериментах, которые могут
продемонстрировать способность дельфинов к решению простых логических задач.
Изучение некоторых форм рассудочной деятельности
Выше, при описании ряда экспериментов с дельфинами, неоднократно шла речь об
их интеллекте, или рассудочной деятельности, по определению Л. В. Крушинского
(1977). В этом разделе мы рассмотрим проблему интеллекта
дельфинов.
Прежде
всего остановимся на интересных работах К. Прайор. В опытах, проведенных на двух
морщинистозубых дельфинах (Steno bredanensis), К. Прайор с соавторами (Pryor et
al., 1969) применили для их обучения метод, ранее не использовавшийся на
животных. В эксперименте вознаграждались не как обычно, отработанные, ранее
сформированные движения дельфина, а только новые, до этого не поощрявшиеся.
Поняв суть задачи, дельфины начинали демонстрировать не только движения,
свойственные их виду, но изобретали новые, не характерные для морщинистозубых
дельфинов.
В
проведенном исследовании авторами было показано, что дельфины могут усвоить
принцип предложенной им задачи, а не только обучиться еще одной новой реакции. В
этом огромное значение проведенной
работы.
Г.
Бейтсон (Bateson, цит. по: Pryor, 1975), присутствовавший при экспериментах,
определил описанное выше поведение у дельфинов как обучение высшего порядка, или
«вторичное обучение». К. Лоренц (1981) оценивает результаты экспериментов с
другой точки зрения. Он считает, что осмысление животными того факта, что
вознаграждаться будет не какое-то определенное движение или система движений, но
любое движение, которое никогда прежде не поощрялось, требует совершенно
неожиданной для животного степени абстрактного
мышления.
К.
Прайор полагает, что с помощью этих опытов ей удалось выявить присущее поведению
высших животных свойство — оригинальность. Это свойство поведения дельфинов
позволяет по-иному подойти к определению существа интеллекта
животных.
Первыми
исследователями, которые поставили проблему оценки умственных способностей у
дельфинов в современной литературе были А. Ф. Мак-Брайд и Д. О. Хебб (McBride,
Hebb, 1948). Всесторонне и подробно проанализировав поведение дельфинов в стаде,
их игры, сексуальные отношения, ориентировочно-исследовательское поведение,
приручение, адаптацию к неволе, авторы пришли к заключению, что с точки зрения
эмоционального поведения и действий, обусловленных внешними причинами, дельфины
по своему умственному развитию занимают промежуточное положение между собакой и
шимпанзе. Хотя авторы и отмечают отсутствие методов, которые могли бы
всесторонне оценить развитие умственных способностей у дельфинов, они указали
путь, который может привести к успеху в изучении этого вопроса. Они полагали,
что нельзя использовать условнорефлекторный критерий для суждения об умственных
способностях животных: не скорость обучения, а решение логических задач может
служить показателем разума у
животных.
Тридцать
лет спустя сходную точку зрения высказал X. Джерисон (Jerison, 1978). По его
мнению, способность к обучению не может служить критерием для изучения
интеллекта у животных, поскольку образование условнорефлекторных связей является
универсальным приспособлением для всех видов животных. Он предлагает разработать
тесты, с помощью которых можно было бы исследовать способность животных к
абстракции.
Исследователями
высказываются различные мнения о возможности познания и изучения интеллекта у
животных. Так, Е. О. Вильсон (Wilson, 1975), Дж. Гарсиа (Garcia, 1978)
подчеркивают, что поведение животных, в том числе и интеллект, следует изучать
только в рамках специфики экологии каждого вида. Авторы полагают, что вряд ли
можно найти такую методику эксперимента, с помощью которой можно было бы
сравнивать интеллект животных, принадлежащих к разным видам. У каждого вида
животных, по их мнению, существует специфический набор поведенческих реакций,
приспособленных к особенностям условий своего существования, необходимых для
выживания.
В то
же время Л. Герман (Herman, 1980), обсуждая выявленные способности дельфинов и
обезьян к оперированию случайными символами, обозначающими предметы и явления
окружающей среды, полагает, что мозг этих животных преадаптирован к высокому
уровню познавательных возможностей. Потенциальные возможности мозга дельфинов и
человекообразных обезьян гораздо выше, чем это необходимо для существования в
естественных условиях. На избыточность потенциальных возможностей мозга
указывали еще А. Н. Северцов (1922), К. Лешли (Lashley, 1949) и Б. Ренш (Rensch,
1960). К. Лешли подчеркивал, что мозг животных, способных к элементарной
абстракции, обладает избыточным резервом нейронов. Л. В. Крушинский (1958),
рассматривая развитие рассудочной деятельности животных, полагает, что эта форма
высшей нервной деятельности не может быть охарактеризована только как частное
приспособление к условиям существования. Рассудочная деятельность, обусловленная
сложнейшей нейронной организацией мозга, развилась как общая адаптивная функция
— арогенез.
Один
из первых экспериментальных подходов к изучению элементарной рассудочной
деятельности у животных в сравнительно физиологическом плане был осуществлен Л.
В. Крушинским и его сотрудниками (см. обзор: Крушинский, 1977). По определению
Л. В. Крушинского, в основе любой, самой сложной формы рассудочной деятельности
животного лежит способность улавливать закономерности, связывающие предметы и
явления окружающей среды, и возможность оперировать этими
закономерностями.
Для
оценки уровня развития рассудочной деятельности у дельфинов были выбраны два
критерия: способность к решению экстраполяционных задач и оценка некоторых
геометрических свойств предметов (оперирование геометрической мерностью
фигур).
Изучение
способности черноморских афалин экстраполировать направление движения исчезающих
за преградой раздражителей при различных условиях их предъявления проводилось в
отсеке размерами 8X10,5 м2, куда была помещена звуко- и светонепроницаемая ширма
длиной 6 м, высотой 2 м, на одну треть погруженная в воду, с окном шириной 0,8 м
в середине (Крушинский и др., 1982). Окно и пространство от ширмы до дна отсека
были забраны сетью. Слева и справа от ширмы имелись проходы шириной около 2 м,
позволявшие дельфину оплывать ширму с любой стороны. При (проведении
эксперимента дельфину, находившемуся на стартовой позиции напротив окна ширмы,
предъявлялся в окне натуральный (рыба) или другой привлекавший его раздражитель
(мяч). После подхода животного к окну по сигналу (всплеску о воду раздражителя)
и при попытке охватить приманку последняя на держателе отодвигалась вдоль ширмы
и скрывалась из поля зрения животного. Таким образом, животному предлагалось
решить задачу отыскания раздражителя: после получения информации о
первоначальном направлении движения раздражителя экстраполировать его
последующий путь за ширмой и обойти ее с соответствующей стороны. В следующем
опыте приманку продвигали в противоположном направлении, а в дальнейшем
направление ее перемещения меняли без строгого
чередования.
Были
проведены 4 серии опытов, различавшихся характером раздражителей и средой, в
которой их предъявляли. В экспериментах первой и второй серий приманкой служила
рыба. В первой серии ее передвигали за ширмой под водой на глубине 0,2 м (22
предъявления). Дельфин правильно решил задачу, допустив только две ошибки. Во
второй серии рыбу 15 раз предъявляли и продвигали за ширмой над водой на высоте
0,2 м. Это не отразилось на решении задачи дельфином, допустившим лишь одну
ошибку.
В
третьей и четвертой сериях опытов, в которых в качестве приманки использовался
мяч, передвигавшийся в 8 пробах по поверхности воды и в 10 — на высоте 0,2 м над
поверхностью, дельфин во всех случаях решил задачу
правильно.
Таким
образом, афалина четко проявила способность к экстраполяции направления
скрывавшихся за преградой как натуральных, так и искусственных раздражителей,
предъявлявшихся под водой и над водой (что исключало возможность их восприятия
за преградой), т. е. показала высокий уровень элементарной рассудочной
деятельности при решении простого варианта экстраполяционной
задачи.
Более
сложный эксперимент, выявляющий способность животных к оперированию мерностью
фигур, основан на том, что из двух фигур, предъявленных животному для выбора —
объемной и плоской, только объемная фигура обладает свойством вмещаемости.
Животные должны понять, что объемная фигура может быть вмещена только в
объемную, но не в плоскую фигуру и в этой фигуре она может быть перемещена из
одной точки пространства в
другую.
Опыты
были проведены с двумя молодыми черноморскими афалинами-самками (Крушинский и
др., 1972). Игру с мячом они часто предпочитали очередному кормлению, и поэтому
в качестве приманки была выбрана их любимая игрушка — мяч.
Рис. 3. Дельфин перед экспериментальной установкой
Фото Л.В.
Крушинского
В эксперименте, который проводили обычно раз в день или через день, одному из
дельфинов предлагалось решить, к какой из двух геометрических фигур (объемной
или плоской) следует подплыть, чтобы найти мяч. Экспериментальная установка
помещалась над водой. На высоте 1 м от поверхности воды укреплялась
горизонтальная штанга, по которой от центра одновременно раздвигались плоская и
объемная пара фигур. В центре установки находился непрозрачный подвижной экран,
поднимая который, можно было скрыть фигуры от дельфинов. К обеим фигурам
прикреплялись педали. Нажимая на них рострумом, дельфин мог повернуть их вокруг
подвески на 180° и из объемной фигуры выпадал мяч. Нажатие на педаль (а в
некоторых опытах — обливание фигур водой изо рта) служило индикатором выбора
дельфином данной фигуры (рис.
3).
Опыт
происходил следующим образом. При открытой ширме дельфину демонстрировали пару
фигур, и в это время экспериментатор играл с ним мячом, привязанным к леске.
Затем поднимали экран, отгораживая от дельфина фигуры, и мячик прятали в
объемную фигуру, отстегивая его от лески. Это занимало 30 — 40 с. Экран
опускали, и, если дельфин был ориентирован строго по центру бассейна, фигуры
раздвигали со скоростью 30 см/с. Дельфин мог подплывать под фигуру и
рассматривать ее спереди из воды и с воздуха. При правильном решении задачи
дельфин подплывал к объемной фигуре, нажимал рострумом на педаль и получал мяч.
В каждом опыте ему предлагали новую пару геометрических фигур. Всего было
использовано 16 пар фигур: девять плоскогранных и семь — тела вращения. Фигуры
были одного цвета, фронтальные проекции которых всегда
совпадали.
Каждая
фигура применялась в эксперименте с одним дельфином не более двух раз с
промежутками между опытами в 3 — 4 недели в целях исключить обучение на фигуры
данной формы.
Так
как фигуры предъявлялись не в воде, а над водой, то необходимой предпосылкой
решения этой задачи являлась способность дельфинов к зрительному различению
геометрических фигур и восприятию глубины
пространства.
Уже
давно известно, что дельфины успешно обучаются зрительному дифференцированию
изображений геометрических фигур как под водой, так и над водой (Kellogg, Race,
1966). Точно рассчитанные прыжки из воды к высоко поднятой над водой цели,
способность точно забрасывать предметы, в сетку свидетельствуют о том, что
дельфины хорошо воспринимают глубину пространства в воздухе. Поэтому задачу,
предложенную нами дельфинам, можно было считать адекватной для их зрительного
восприятия.
Результаты
опытов с афалинами показали, что оба животных с первого раза правильно решали
задачу с геометрической мерностью фигур. Особенно интересны были два первые
опыта с одной из самок по кличке Василиса, когда фигуры не имели педалей. После
раздвижения фигур в обоях случаях Василиса подходила к объемной фигуре и
обливала ее водой — реакция выпрашивания мяча. Поскольку афалина не получала в
этих опытах мяч, то в последующих опытах она просто уходила от экспериментальной
установки. Впоследствии во всех экспериментах к фигурам стали прикреплять
педали.
Суммарные
данные опытов, проведенных с двумя афалинами, продемонстрировали явное
преобладание правильных решений (из 48 опытов — 37 правильных выборов,
р<0,001).
Следующие
доводы, с нашей точки зрения, исключают возможность объяснения выбора дельфинами
объемных фигур в результате обучения. Во-первых, каждый опыт проводился с
разными по форме фигурами. Во-вторых, два первые опыта с Василисой, когда фигуры
были без педалей, показали, что оба раза она правильно решила задачу, не получая
мяча. В-третьих, неправильные решения задачи не были приурочены к началу опытов,
как это обычно бывает в опытах по обучению дифференцированию геометрических
фигур (Kellogg, Rice, 1966). Ошибки в экспериментах с мерностью фигур равномерно
распределялись по всей серии опытов. Появление ошибочных выборов плоской фигуры
у обеих афалин совпадало с перевозбуждением животных в связи с разными
причинами.
Проведенные
эксперименты с афалинами показали, что они способны без предварительного
обучения строить программу поведения, в основе которой лежит способность к
оперированию мерностью фигур, т.е. они имеют представление об элементарных
геометрических свойствах предмета — вмещаемости объемных фигур.
Рис. 4. Индивидуальные кривые накопления правильных ответов
А —
при решении задачи оперирования эмпирической мерностью фигур афалиной Василисой;
Б — во время выработки дифференцирования геометрических фигур у афалины Леди. 1
— правильный ответ; 2 — неправильный ответ. По оси ординат — разность между
числом правильных и неправильных ответов; по оси абсцисс — Число (N)
предъявлений задачи
Описанную выше задачу предлагали другим животным: обезьянам
(Cercopithecidae), собакам, кошкам и морским львам (Zalophus californianus). В
результате проведенных опытов (Крушинский и др., 1972; Дашевский, 1972;
Крушинский и др., 1976; Крушинская, Дмитриева, 1974) было показано, что только
обезьяны и афалины способны к ее решению без специального обучения. В процессе
многочисленных предъявлений задачи у собак удалось выработать дифференцирование
объемных фигур от плоских. Однако, как показали контрольные эксперименты, даже в
результате длительного обучения собаки оказались неспособными к улавливанию
(абстракции) свойства вмещаемости объемных фигур, они научились лишь различать
объемные и плоские фигуры (Крушинский и др.,
1976).
Приведенные
исследования дают основание считать, что афалины способны решать элементарные
логические задачи, требующие экстраполяции направления движений раздражителей и
оперирования эмпирической мерностью фигур. Это свидетельствует о наличии у этих
животных способности к абстракции. Следует также отметить, что эксперименты были
построены не на традиционном пищевом подкреплении, а на игровом. Результаты
опытов подтверждают мнение целого ряда исследователей, что дельфины (афалины)
обладают высокоразвитым интеллектом, сопоставимым с таковым у обезьян.
Заключение
Приведенные в настоящей главе данные свидетельствуют о чрезвычайно высоком
уровне развития мозга китообразных и весьма пластичных и сложных формах их
поведения. Мощная теленцефализация китообразных не может, по-видимому, быть
объяснена лишь высокой специализацией этих животных к водному образу жизни и
развитой у них способностью к эхолокации, как полагают некоторые исследователи
(Кесарев, 1974; Колдуэлл, Колдуэлл, 1978; Томилин, 1980), а связана в первую
очередь с развитием их высших психических функций: сложных форм обучения и
рассудочной
деятельности.
Исследование
высшей нервной деятельности дельфинов методом условных рефлексов показало, что
по основным видам условнорефлекторных связей дельфины существенно не отличаются
от других высших наземных позвоночных. Это подтверждает известное положение в
теории обучения, согласно которому в основе приобретения индивидуальных навыков
у всех позвоночных лежат одни и те же процессы (Skinner, 1956; Воронин,
1969).
Переделка
сигнального значения раздражителей при их дифференцировании рассматривается
(Bitterman, 1965) одним из показателей качественного различия способности к
обучению у позвоночных животных, обладающих разным уровнем организации нервной
системы. Однако применение этой методики для оценки познавательных способностей
дельфинов не выявило у них существенных отличий от других
позвоночных.
Изучение
более сложных форм поведения, таких, как имитационное научение, показало, что
своего наивысшего развития оно достигает у дельфинов и шимпанзе, проявляясь в
подражании орудийной деятельности человека. Подражание различным формам
орудийной деятельности человека, проявляющееся в виде игр и сбора пищевых
объектов, могут служить одним из убедительных доводов в пользу высокого уровня
их рассудочной деятельности. Имитационное поведение дельфинов является
предпосылкой для возникновения в сообществах китообразных определенных традиций,
которые передаются от поколения к
поколению.
Способность
к имитационному обучению и образование условнорефлекторных реакций на тождество
сигналов (выбор по образцу) у дельфинов должна рассматриваться как выражение
функций сложного обобщения. Исследование возможности выработки у дельфинов
наличного и отсроченного выбора по тождеству сигналов выявило у них способность
к решению этих задач того же уровня, что и у обезьян. Процесс образования выбора
по тождеству сигналов осуществляется животными не на основе жесткой системы
«сигнал — реакция», а путем обобщения одного или нескольких признаков
раздражителя. Такая форма обобщений у животных классифицируется исследователями
как «интеллектуальные формы
поведения».
Способность
дельфинов обучаться отсроченному выбору звукового стимула-образца среди
нескольких тестовых сигналов является показателем высокого уровня лабильности их
оперативной памяти и надежности в реализации следовых процессов. Уровень
развития оперативной памяти у дельфинов можно рассматривать как одну из
предпосылок к отвлеченной символизации предметов и явлений окружающей их
среды.
Многочисленные
исследования, направленные на изучение «языковых» способностей дельфинов, до сих
пор не подтвердили гипотезы о существовании у дельфинов особых способов
коммуникаций, принципиально отличающихся от сигнализаций других животных и тем
более тех, по которым их можно было бы приблизить к языковому общению
человека.
В
настоящее время исследуются возможности усвоения дельфинами звуковых
символов-признаков предметов и явлений окружающего их мира, тех качеств, которые
лежат в основе человеческого языка. Выяснена возможность обучения дельфинов
реагировать определенными действиями на различные комбинации случайно выбранных
звуков, что рассматривается как первый этап на пути к установлению двусторонней
связи человека с дельфином на основе использования языковых
символов.
Как
известно, такой путь исследований привел к успешным результатам при обучении
шимпанзе азбуке жестов (ASL). Обученные по системе ASL шимпанзе могли
использовать эти знаки-символы в качестве средства общения с экспериментаторами
и другими
обезьянами.
Возможно
ли будет осуществить полностью намеченную программу исследований по установлению
двусторонней связи человека и дельфинов, зависит от ряда
причин.
Успех
исследований в большой мере зависит от адекватности выбранной формы эксперимента
физиологическим возможностям воспринимающих и эффекторных систем дельфина.
Высокий уровень развития слухового анализатора дельфинов и способности к
имитации искусственных звуковых сигналов широкого диапазона могут служить
залогом успешного проведения запланированной программы
исследования.
Вместе
с тем нельзя исключить из рассмотрения возможность существования у дельфинов
систем кодирования информации, принципиально отличающихся от привычных
представлений человека. Например, сложная информация может кодироваться не
последовательностью символов во времени, как принято в языке человека (и,
видимо, в системах общения у обезьян), а одновременной передачей набора
составляющих частотного спектра звуков или каким-либо другим необычным для
человека и пока нераскрытым способом. Очевидно, в этом случае навязываемая
дельфину экспериментатором «человеческая» система символов может для него
оказаться совершенно
непривычной.
И,
наконец, возможность установления двусторонней связи человека и дельфина на
основе «языковой» символизации зависит от того, обладают ли дельфины уровнем
развития умственных способностей, необходимым для оперирования отвлеченными
символами.
Исследование
рассудочной деятельности у дельфинов на основе решения ими простых логических
задач (экстраполяция направления движения раздражителя и оперирование
имперической размерностью фигур) можно рассматривать как один из этапов подхода
к познанию интеллекта этих животных. Экспериментально было показано, что
дельфины и обезьяны способны к решению наиболее сложных задач, требующих
соизмерения пространственных свойств объемных и плоских предметов, и на
основании уловленных при этом закономерностей строить адекватное поведение в
новой ситуации. Рассматривая предложенные тесты как объективные параметры для
оценки рассудочной деятельности животных, можно заключить, что дельфины и по
этому критерию стоят на том же уровне, что и
обезьяны.
Обобщая
данные морфологических исследований и экспериментального изучения поведения
китообразных, можно сказать, что они в совокупности однозначно свидетельствуют о
больших потенциальных возможностях их мозга для оперирования большим объемом
отвлеченных символов.