АКСОНЫ: Механизмы выбора пути
АКСОНЫ: Механизмы выбора пути
Со времени публикации была проделана работа по развитию прототипа системы индивидуальной адаптивности Beast (fornit.ru/beast) и появилось много новых данных и материалов с которыми можно ознакомисться в трилогии «Основы фундаментальной теории сознания» и точки входа теории МВАП «Организации механизмов мозга человека».
AXON GUIDANCE MECHANISMS AND MOLECULES: LESSONS FROM
INVERTEBRATES
Sofia J. Araujo & Guy Tear
Nature Reviews Neuroscience 4, № 11. P. 910 -922
(2003)
Перевод И.Г. Лильп (lilp@mail.ru)Nature Reviews Neuroscience 4, № 11. P. 910 -922 (2003)
Механизмы установления межнейронных связей в нервной системе позвоночных и беспозвоночных являются в настоящее время одной из важных проблем в области нейробиологии. Исследователи, занимающиеся изучением этого процесса, приступили к идентификации молекул и механизмов, лежащих в основе этого процесса. Как модельные системы каждая группа организмов имеет свои преимущества и вносит определенный вклад в понимание того, каким образом аксоны достигают своих конечных мишеней. Именно у беспозвоночных модельных организмов при использовании классических методов скрининга мутаций были обнаружены гены, нарушающие процесс нахождения аксонами своего пути во время развития мозга. Позвоночные организмы являются хорошими модельными системами для изучения функциональных механизмов. В настоящем обзоре авторы рассматривают роль беспозвоночных животных в открытии молекулярных механизмов, управляющих направлением движения аксонов к своим конечным мишеням.
Аксоны часто растут вдоль др. аксонов, чтобы создать пучок, называемый fascicles, и целый ряд cell adhesion molecules (CAMs) обнаруживается на поверхности аксонов, которые вносят вклад в процесс фасцикуляции.
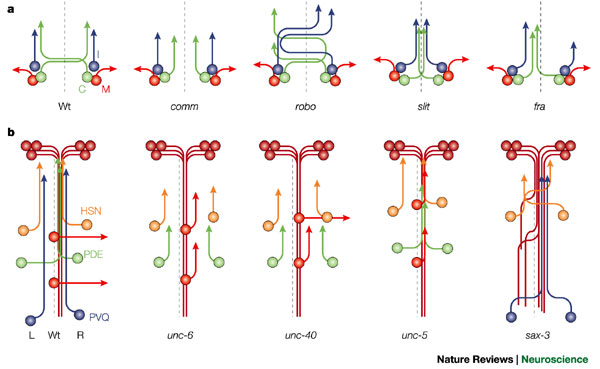 Axon pathways at the CNS midline of Drosophila and Caenorhabditis elegans. a Simplified schematic of the trajectories taken by commissural (C, green), ipsilateral (I, blue) and motor (M, red) neurons in the Drosophila central nervous system (CNS). In the wild-type (Wt) embryo, most CNS axons extend along a commissural pathway and cross the midline (dashed line) in one of two commissural axon tracts. These axons cross the midline only once. The ipsilaterally projecting axons extend on one side of the CNS only, whereas the motor neurons extend out to the periphery either on their own side of the CNS or after crossing the midline (not shown). The Drosophila CNS is bilaterally symmetrical, with the same organization of neurons on either side of the midline. In commissureless (comm) mutants, the C neurons fail to cross, whereas in the robo mutant, I axons can cross the midline and C axons can recross, resulting in whorls of axons at the midline. In the absence of slit, all CNS axons extend towards the midline region and are unable to leave. When frazzled (fra) (or both netrin genes) is removed some axons fail to cross the midline, and breaks appear in the longitudinal tracts. b | Simplified schematic of the trajectories of some neurons in the C. elegans ventral nerve cord (VNC). The C. elegans VNC is asymmetric, with many more axons running in the right fascicle (R) than in the left fascicle (L). Many axons enter the VNC from the anterior or posterior end of the tract; for example, the AV (brown) or PVQ (blue) neurons. Other axons join the VNC along its length; for example, the HSN (orange) and PDE (green) axons, and they might extend contralaterally (PDEL) or ipsilaterally (PDER) once in the VNC. Motor neurons, such as DA5 and DA6 (red), have their cell bodies at the ventral midline (dashed line), and they extend dorsally away from the VNC. In unc-6 mutants, the D neurons fail to extend away from the VNC, and the PDE and HSN neurons fail to extend to the VNC. Mutations in unc-40 primarily affect the axons extending to the VNC, but they can also disrupt extension away from the ventral midline. In the absence of unc-5 activity, the D motor neurons are unable to extend dorsally. Mutations in sax-3 cause a VNC phenotype, where axons from the AV, HSN or PVQ neurons fail to remain in their fascicle and inappropriately cross the midline.
Axon pathways at the CNS midline of Drosophila and Caenorhabditis elegans. a Simplified schematic of the trajectories taken by commissural (C, green), ipsilateral (I, blue) and motor (M, red) neurons in the Drosophila central nervous system (CNS). In the wild-type (Wt) embryo, most CNS axons extend along a commissural pathway and cross the midline (dashed line) in one of two commissural axon tracts. These axons cross the midline only once. The ipsilaterally projecting axons extend on one side of the CNS only, whereas the motor neurons extend out to the periphery either on their own side of the CNS or after crossing the midline (not shown). The Drosophila CNS is bilaterally symmetrical, with the same organization of neurons on either side of the midline. In commissureless (comm) mutants, the C neurons fail to cross, whereas in the robo mutant, I axons can cross the midline and C axons can recross, resulting in whorls of axons at the midline. In the absence of slit, all CNS axons extend towards the midline region and are unable to leave. When frazzled (fra) (or both netrin genes) is removed some axons fail to cross the midline, and breaks appear in the longitudinal tracts. b | Simplified schematic of the trajectories of some neurons in the C. elegans ventral nerve cord (VNC). The C. elegans VNC is asymmetric, with many more axons running in the right fascicle (R) than in the left fascicle (L). Many axons enter the VNC from the anterior or posterior end of the tract; for example, the AV (brown) or PVQ (blue) neurons. Other axons join the VNC along its length; for example, the HSN (orange) and PDE (green) axons, and they might extend contralaterally (PDEL) or ipsilaterally (PDER) once in the VNC. Motor neurons, such as DA5 and DA6 (red), have their cell bodies at the ventral midline (dashed line), and they extend dorsally away from the VNC. In unc-6 mutants, the D neurons fail to extend away from the VNC, and the PDE and HSN neurons fail to extend to the VNC. Mutations in unc-40 primarily affect the axons extending to the VNC, but they can also disrupt extension away from the ventral midline. In the absence of unc-5 activity, the D motor neurons are unable to extend dorsally. Mutations in sax-3 cause a VNC phenotype, where axons from the AV, HSN or PVQ neurons fail to remain in their fascicle and inappropriately cross the midline. Во время эмбрионального развития аксоны должны преодолеть значительные расстояния, чтобы достичь своих конечных мишеней. Пониманию этого процесса в значительной степени способствовали исследования на беспозвоночных моделях. Такие организмы как Drosophila и C. elegans оказались особенно полезными из-за их относительно просто устроенной нервной системы и возможности проведения точного генетического анализа.
Ключевой стратегией во время исследования нахождения аксонами своего пути является нарушение пути аксона к мишени между клетками-ориентирами (guidepost), существование которых было обнаружено в почке (зачатке) конечности кузнечика. Важными посредниками во взаимодействии между клетками-ориентирами и аксонами являются нетрин (netrin) и его рецепторы. Генетические исследования позволили очертить контуры сигнального каскада, опосредующего действие нетрина. Roundabout (Robo) и Slit формируют часть связанной между собой сигнальной системы, являющейся критической в процессе нахождения аксоном своего пути. Их главная роль заключается в регулировании (упорядочивании) средней линии (midline), пересекаемой аксонами в нервной системе Drosophila. Серия генетических исследований позволила выявить каскад, который может опосредовать их эффект.
Уже имеющиеся (pre-existing) аксоны важны для выбора направления движения и роста последующих аксонов, поскольку ведомые аксоны часто fasciculate (расположены в виде пучка) с направляющим аксоном, который достигает своей мишени. Предполагают, что траектории аксонов имеют разные молекулярные метки, распознаваемые последующими аксонами. Было идентифицировано несколько молекул, выполняющих роль меток, и среди них молекулы neuroglian, N-cadherin и fasciclins.
Defasciculation также важна, как и fasciculation, для нахождения аксоном пути к своей мишени. Уже идентифицировано несколько молекул, участвующих в этом процессе – среди них semaphorins, Beaten path, Off-track и другие. Внутриклеточные пути, посредством которых эти молекулы опосредуют defasciculation, изучаются. Исследование глаза Drosophila позволило идентифицировать дополнительные молекулы-проводники, которые не были идентифицированы при использовании обычного фенотипического скрининга. Особенно это было полезно для идентификации внутриклеточных сигнальных элементов, которые участвуют в нахождении аксонами своего пути.
Несмотря на идентификацию многих молекул, лежащих в основе «управления» аксонами, вероятно, существуют и другие молекулы. Например, мы мало знаем о том, как аксоны интегрируют полученные ими одновременно сигналы и генерируют единственный верный ответ, а также как они адаптируют свои ответные реакции, двигаясь вдоль своего пути следования к конечной мишени. Чтобы ответить на эти вопросы, необходимо использовать преимущества как беспозвоночных, так и позвоночных модельных систем.
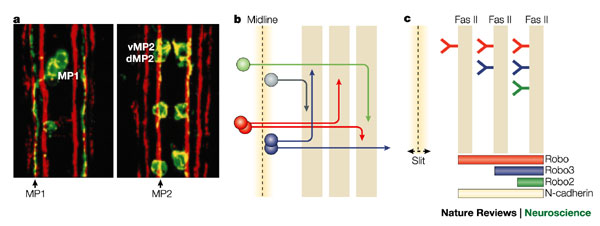 Axon pathways in the Drosophila CNS. a Fasciclin II labels three main fascicles on either side of the Drosophila midline. Stage 17 embryos, showing the medial, intermediate and lateral FasII fascicles in red and pioneer axons of MP1, dMP2 and vMP2 neurons in green. At this stage, axons of dMP2 and vMP2 run along the first (medial) FasII fascicle, and axons of the MP1 run along the second (intermediate) FasII fascicle. Axons of pCC also run along the medial FasII fascicle (not shown). According to these observations, the medial FasII-positive tract has been named the pCC/MP2 pathway, and the intermediate tract the MP1 pathway89. b | Schematic representation of some axonal pathways joining the various longitudinal fascicles parallel to the midline. Each follower axon can specifically recognize the surface of particular axons that have extended earlier. These axons can select to join pre-existing pathways and bundle together to form the fascicles. The axons switch between these pathways as they extend towards their own individual target. c | The Robo code model of how axons choose their pathway on either side of the midline. The Slit protein is secreted by midline cells and diffuses to create a gradient across the central nervous system. The distance travelled by the axons away from the midline depends on the combination of Robo proteins that they express. Axons that express Robo protein continuously do not cross the midline, but choose a longitudinal fascicle on their own side according to which other Robo proteins they express. Axons that do not initially express the Robo protein cross the midline, where Robo levels now increase, and they choose a FasII-positive pathway depending on the combination of Robo proteins that they now express. Axons expressing Robo, Robo2 and Robo3 are most strongly repelled from the midline and enter the lateral fascicle. Axons expressing only Robo and Robo3 extend into the intermediate zone and axons that only express Robo stay closer to the midline in the medial zone. As the level of Slit signalling through the Robo receptors decreases, the neurons might recognize N-cadherin and remain in their correct zone. The axons then use their specific surface markers (such as FasII or Connectin) to select the appropriate pathway. Reproduced, with permission, from Ref. 89 © Company of Biologists (1997).
Axon pathways in the Drosophila CNS. a Fasciclin II labels three main fascicles on either side of the Drosophila midline. Stage 17 embryos, showing the medial, intermediate and lateral FasII fascicles in red and pioneer axons of MP1, dMP2 and vMP2 neurons in green. At this stage, axons of dMP2 and vMP2 run along the first (medial) FasII fascicle, and axons of the MP1 run along the second (intermediate) FasII fascicle. Axons of pCC also run along the medial FasII fascicle (not shown). According to these observations, the medial FasII-positive tract has been named the pCC/MP2 pathway, and the intermediate tract the MP1 pathway89. b | Schematic representation of some axonal pathways joining the various longitudinal fascicles parallel to the midline. Each follower axon can specifically recognize the surface of particular axons that have extended earlier. These axons can select to join pre-existing pathways and bundle together to form the fascicles. The axons switch between these pathways as they extend towards their own individual target. c | The Robo code model of how axons choose their pathway on either side of the midline. The Slit protein is secreted by midline cells and diffuses to create a gradient across the central nervous system. The distance travelled by the axons away from the midline depends on the combination of Robo proteins that they express. Axons that express Robo protein continuously do not cross the midline, but choose a longitudinal fascicle on their own side according to which other Robo proteins they express. Axons that do not initially express the Robo protein cross the midline, where Robo levels now increase, and they choose a FasII-positive pathway depending on the combination of Robo proteins that they now express. Axons expressing Robo, Robo2 and Robo3 are most strongly repelled from the midline and enter the lateral fascicle. Axons expressing only Robo and Robo3 extend into the intermediate zone and axons that only express Robo stay closer to the midline in the medial zone. As the level of Slit signalling through the Robo receptors decreases, the neurons might recognize N-cadherin and remain in their correct zone. The axons then use their specific surface markers (such as FasII or Connectin) to select the appropriate pathway. Reproduced, with permission, from Ref. 89 © Company of Biologists (1997).
 Motor neuron axon pathways in Drosophila. a | Abdominal motor axon projections in the wild type and in a Dlar mutant. Wild-type and Dlar motor neuron branches as seen in filleted stage 17 embryos. The intersegmental nerve b (ISNb) defasciculates from the ISN and innervates ventral longitudinal muscles 6, 7, 12 and 13. In Dlar mutants, the ISNb fails to defasciculate from the ISN (arrowhead) and does not innervate the ventral longitudinal muscles126. The asterisk marks the position where ISN and ISNb can be distinguished as separate fascicles. ISNb, intersegmental nerve b. Reproduced, with permission, from Ref. 123 © Elsevier Sciences (1996). b | Schematic diagram of wild-type motor axon projections. Each main nerve branch is shown in a different colour as it emerges from the exit junction outside the central nervous system (CNS). In wild-type motor axons, the ISN (red) and SN (orange/green) defasciculate into five pathways that innervate the muscle fibres. The SNa (green) and SNc (orange) pathways emerge from the SN root and the ISN (red), ISNb (blue) and ISNd (brown) pathways arise from the ISN root. c | side mutants display an extreme bypass phenotype. In side mutants, as in beat and Dlar mutants, axons from the main motor axon pathways fail to defasciculate at choice points and do not innervate most of the ventral musculature.
Motor neuron axon pathways in Drosophila. a | Abdominal motor axon projections in the wild type and in a Dlar mutant. Wild-type and Dlar motor neuron branches as seen in filleted stage 17 embryos. The intersegmental nerve b (ISNb) defasciculates from the ISN and innervates ventral longitudinal muscles 6, 7, 12 and 13. In Dlar mutants, the ISNb fails to defasciculate from the ISN (arrowhead) and does not innervate the ventral longitudinal muscles126. The asterisk marks the position where ISN and ISNb can be distinguished as separate fascicles. ISNb, intersegmental nerve b. Reproduced, with permission, from Ref. 123 © Elsevier Sciences (1996). b | Schematic diagram of wild-type motor axon projections. Each main nerve branch is shown in a different colour as it emerges from the exit junction outside the central nervous system (CNS). In wild-type motor axons, the ISN (red) and SN (orange/green) defasciculate into five pathways that innervate the muscle fibres. The SNa (green) and SNc (orange) pathways emerge from the SN root and the ISN (red), ISNb (blue) and ISNd (brown) pathways arise from the ISN root. c | side mutants display an extreme bypass phenotype. In side mutants, as in beat and Dlar mutants, axons from the main motor axon pathways fail to defasciculate at choice points and do not innervate most of the ventral musculature.
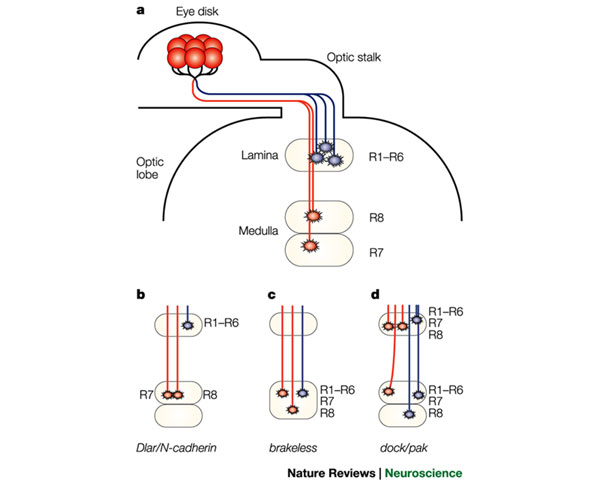 Projections of R-cell axons to targets in the optic lobe. a | A single ommatidium containing eight R-cell neurons is shown. R-cell axons project through the optic stalk into the optic lobe, where they contact their targets. R1–R6 axons (blue) stop at their target layer in the lamina. The R7 and R8 axons (red) continue into the underlying medulla, where they stop in two distinct layers. b–d | Schematic representations of various R-cell mutant phenotypes. b | In Dlar and N-cadherin mutants R7 and R8 terminate in the R8 region. c | In the absence of Brakeless, R1–R8 all terminate in the medulla. d | In dock and pak mutants, uneven axon bundles exit the optic stalk towards the lamina. Most fibres follow abnormal paths, with R1–R6 failing to terminate in the lamina and innervating the medulla. As a result, some regions are hyperinnervated, whereas others remain uninnervated.
Projections of R-cell axons to targets in the optic lobe. a | A single ommatidium containing eight R-cell neurons is shown. R-cell axons project through the optic stalk into the optic lobe, where they contact their targets. R1–R6 axons (blue) stop at their target layer in the lamina. The R7 and R8 axons (red) continue into the underlying medulla, where they stop in two distinct layers. b–d | Schematic representations of various R-cell mutant phenotypes. b | In Dlar and N-cadherin mutants R7 and R8 terminate in the R8 region. c | In the absence of Brakeless, R1–R8 all terminate in the medulla. d | In dock and pak mutants, uneven axon bundles exit the optic stalk towards the lamina. Most fibres follow abnormal paths, with R1–R6 failing to terminate in the lamina and innervating the medulla. As a result, some regions are hyperinnervated, whereas others remain uninnervated.