Системные основы интеллекта
Системные основы интеллекта
К.В.Судаков
НИИ нормальной физиологии им. П.К.Анохина РАМН
Аннотация
В статье приведены материалы, свидетельствующие о новых аспектах понимания функций мозга на основе теории функциональных систем, постулированной П.К.Анохиным.
В отличие от распространенной проводниковой парадигмы организации функций мозга, в статье рассматриваются корково-подкорковые взаимоотношения, обусловленные деятельностью множества функциональных систем – организма. Показано, что в каждый данный момент времени деятельность мозга строится доминирующей мотивацией. Специально рассмотрен вопрос о голографическом механизме взаимодействия мотивации и подкрепления на структурах мозга.
Постулируется положение о том, что доминирующие мотивации выступают в качестве структурно-функциональной “канвы”, на которой подкрепляющие возбуждения формируют “отпечатки действительности” – динамические стереотипы. Рассматривается гипотеза о роли аппарата акцептора результата действия в качестве голографического экрана функциональных систем. Обогащение акцептора результата действия в течение индивидуальной жизни формирует интеллект субъектов, а оценка акцептором результата действия разнообразных параметров действительности – сознание.
Интеллект в общем смысле слова – это сгусток жизненного опыта каждого индивида и, в значительной степени, – всех его предшествующих поколений.
К.Ясперс (1997) обозначает интеллект как совокупный умственный потенциал человека, включающий многочисленные врожденные и приобретенные способности, которые он целесообразно использует для адаптации к различным условиям жизни.
Современная наука о мозге еще не располагает достаточными сведениями, позволяющими раскрыть свойства интеллекта человека. В сознании современных нейрофизиологов и нейропсихологов неизменно господствует рефлекторный подход к функциям мозга. А это определяет “проводниковую” манеру мышления, когда процесс воздействия того или иного фактора на организм рассматривается как проекция соответствующих возбуждений в определенные зоны мозга, а эффекторные функции – как результат первичного возбуждения специальных исполнительных центров и дальнейшего распространения центральных возбуждений на исполнительные органы. В результате мозг представляется как совокупность сенсорных проекционных полей, взаимодействующих с моторными и вегетативными центрами.
Несколько изменяет эти представления химическая интеграция мозговых структур, т.к. показано, что адренергические, холинергические и серотонинергические нейроны широко представлены в различных подкорковых образованиях и в коре мозга (Robbins T.W. et al., 1992). Однако это не изменило традиционной “проводниковой” парадигмы построения функций мозга.
В “Рефлексах головного мозга” И.М.Сеченов постулировал широко известную фразу: “Все акты сознательной и бессознательной деятельности по способу происхождения суть рефлексы”.
Обратим, однако, внимание на другую фразу И.М.Сеченова: “Предметный мир существовал и будет существовать, по отношению к каждому человеку, раньше его мысли; следовательно, первичным фактором в развитии последней всегда был и будет для нас внешний мир с его предметными связями и отношениями” (Сеченов И.М., 1947).
Именно внешний мир определяет по И.М.Сеченову деятельность головного мозга человека.
Тем не менее, вопрос о том, как осуществляется формирование функций мозга, интеллекта и сознания под воздействием окружающей субъектов действительности, остается открытым до настоящего времени.
Теория функциональных систем, сформулированная П.К.Анохиным (Анохин П.К., 1978, 1998), позволила по-новому взглянуть на организацию функций мозга. В отличие от традиционных представлений, связывающих функции мозга с действием специальных стимулов, порождавших отраженные ответные реакции организма, теория функциональных систем раскрывает внутренние механизмы деятельности мозга, которые определяют целенаправленное поведение и психическую деятельность человека и животных.
Согласно теории функциональных систем целый организм представляет гармоническое взаимодействие множества функциональных систем различного уровня: молекулярного, гомеостатического, поведенческого и популяционного (Судаков К.В., 2000).
Центральные звенья многих функциональных систем охватывают мозговые структуры различного уровня (Рис. 1). Взаимодействие разных функциональных систем на уровне головного мозга всегда строится по принципу доминирования, открытого А.А.Ухтомским (1925). В каждый данный момент времени деятельностью мозга завладевает ведущая для сохранения жизни или адаптации организма во внешней среде функциональная система. Каждую такую систему характеризует некое общее эмоциональное состояние организма, обеспечивающие АО-первых, наиболее оптимальные реакции и специализацию, и во-вторых - личностное отношение и мотивацию ответных реакций. Другие функциональные системы в это время или способствуют деятельности доминирующей функциональной системы, или вытормаживаются. После удовлетворения потребности организма, сформировавшей доминирующую функциональную систему, начинает доминировать другая ведущая по социальной и биологической значимости функциональная система и т.д.
Динамика формирования функциональных систем во времени следует принципу системного квантования процессов жизнедеятельности (Судаков К.В., 1997). Согласно этому принципу, непрерывный континуум процессов жизнедеятельности расчленяется в живых организмах на дискретные отрезки – “системокванты” от потребности к ее удовлетворению.
Каждый “системоквант” включает формирование соответствующей потребности, мотивацию и поведение, направленное на удовлетворение этой потребности, а также постоянную оценку с помощью обратной афферентации достигнутых промежуточных и конечного результатов, удовлетворяющих эту потребность. Мозговая архитектоника, обеспечивающая “системокванты”, включает все системные компоненты, установленные П.К.Анохиным: афферентный синтез, принятие решения, акцептор результата действия и оценку параметров достигнутых результатов акцептором результата действия на основе поступающей в мозг обратной афферентации.
Ведущими критическими состояниями “системоквантов” поведенческой и психической деятельности человека и животных являются мотивация и подкрепление, как адаптивный результат, удовлетворявший лежащую в основе доминирующей мотивации потребность.
В механизмах доминирования функциональных систем в мозге ведущая роль принадлежит биологическим и социальным мотивациям.
Биологические мотивации, как показали наши исследования (Судаков К.В., 1992), строятся по пейсмекерному принципу. Пейсмекеры (самопроизвольно генерирующие импульсы нейроны) биологических мотиваций локализованы в гипоталамических структурах мозга. Благодаря обширным морфофункциональным связям с другими структурами мозга и с корой больших полушарий, гипоталамические пейсмекерные нейроны держат практически весь мозг в зависимости от их состояния.
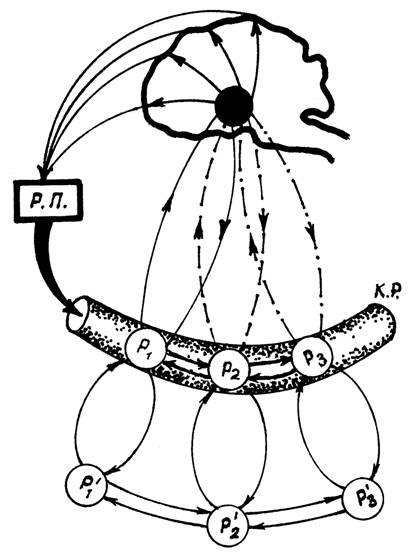
Рис. 1. Схематическое изображение взаимодействия множества функциональных систем на структурах мозга. На схеме показано, что ведущая потребность формирует доминирующую мотивацию, завладевавшую деятельностью мозга и организующую поведение. Р1', Р2', Р3' – результаты деятельности функциональных систем метаболического, а Р1, Р2, Р3 – гомеостатического уровня. РП – результат поведения, КР – кровяное русло.
Социальные мотивации, формирующиеся исходно на основе биологических мотиваций, строятся также по пейсмекерному принципу. При этом пейсмекерные нейроны, определявшие тоническую активность социальных мотиваций, локализуются, как установили (Леонтьев А.Н., 1976; Урываев Ю.В., 1978), в различных, преимущественно в лобных отделах коры больших полушарий и интегрируют возбуждения разнообразных сенсорных модальностей, поступающие в соответствующие проекционные зоны коры мозга.
Как показали наши опыты (Судаков К.В., 1996) и опыты других исследователей (Громова Е.А., 1960), доминирующие биологические мотивации голода, жажды и страха отчетливо проявляются в генерализованной по структурам гипоталамуса, таламуса, лимбических образований мозга и в коре больших полушарий реакции ЭЭГ-активации.
Восходящие активирующие влияния мотивациогенных центров на другие структуры мозга и кору больших полушарий составляют “жизненную энергию каждого из нас” (Павлов И.П., 1951. – С. 199).
Доминирующие мотивации тесно связаны с другой ведущей стадией системной архитектоники поведенческих актов – акцептора результата действия, составлявшего направлявший компонент поведения (Судаков К.В., 1998). Корковые пейсмекеры, создаваемые на основе процессов афферентного синтеза, особенно пейсмекеры лобных отделов больших полушарий, обладают особыми свойствами. Они, участвуя в формировании предпусковой интеграции, определяют, как показал Ю.В.Урываев (1978), выбор доминирующей биологической мотивации. При этом они оказывают специфические нисходящие тормозные и активирующие влияния на пейсмекеры биологических мотиваций, расположенные в гипоталамической области (Зилов В.Г., 1965).
С другой стороны, пейсмекеры лобной коры определяют дистантную синхронизацию ЭЭГ височной и затылочной коры и осуществляют настройку определенных анализаторов на восприятие сигнальных раздражителей, определяя тем самым межсигнальное взаимодействие анализаторов (Урываев Ю.В., 1978).
Исследования сотрудников нашего Института показали, что в организации мотивационных состояний, определяемых активностью того или иного пейсмекера, существенная роль, наряду с нейромедиаторами, принадлежит олигопептидам. Одни из этих эндогенных олигопептидов усиливают мотивационные состояния, другие, наоборот, тормозят.
Стимулирующим пищевые мотивации действием обладают окситоцин, бета-липотропин, пентагастрин, а оборонительные мотивации – брадикинин. Ангиотензин-II, аргинин-вазопрессин, пептид, вызывавший дельта-сон, вещество П, бета-эндорфин и другие олигопептиды при введении кроликам в боковые желудочки мозга подавляли оборонительные и пищевые мотивации (Судаков К.В., 1988).
Вазопрессин и окситоцин, как показал С.К.Судаков (1987), определяют доминирование оборонительной мотивации.
В исследованиях (Судаков К.В. с соавт., 1987) показано, что при введении олигопептидов в боковые желудочки мозга изменяется модальность поведения при раздражении мотивациогенных центров гипоталамуса. Можно думать, что при этом реорганизуется корково-подкорковая архитектоника доминирующих биологических мотиваций.
Содержание олигопептидов, таких как вещество П, пептида, вызывавшего дельта-сон, ангиотензина-II и других в гипоталамусе определяет устойчивость животных к эмоциональному стрессу (Судаков К.В., 1998) и к формированию алкогольной мотивации (Судаков К.В., 1994).
В организации cистемных функций мозга ведущую роль играют взаимодействия доминирующих мотиваций с соответствующим подкреплением, т.е. с удовлетворением лежащих в их основе потребностей. Взаимодействие мотивационных и подкреплявших возбуждений на нейронах мозга строится по голографическому принципу (Судаков К.В., 1997). Нервная и гуморальная сигнализация от потребности выступает по аналогии с физической голографией в роли опорной волны. Сигнализация от различных параметров подкрепления выступает в качестве предметной волны. При этом нейроны различных отделов мозга, включенные в доминирующее мотивационное состояние, в паттернах межимпульсных интервалов своей разрядной деятельности специфически отражают соответствующую потребность. Модное название оказалось излишне привлекательным для автора. Конечно, оно здесь особенно не вредит, но и ничего нового и не вносит, просто уводя логику рассуждения в бесперспективное русло. Процентное содержание таких нейронов, участвующих в организации биологических мотиваций, оказалось выше в стволовых структурах мозга и убывает по направлению к коре больших полушарий: их больше в стволовых образованиях ретикулярной формации таламуса и гипоталамуса (Рис. 2).
Подкрепление переводит пачкообразную активность этих нейронов в регулярную. Характерно, что по мере неоднократных подкреплений, например, при выработке условных рефлексов, регулярная активность у нейронов, исходно включенных в доминирующую мотивацию, начинает опережающе проявляться на действие условных сигналов и даже на обстановку эксперимента (Журавлев Б.В., 1986).
Голографический принцип построения доминирующих мотиваций и их взаимодействия на нейронах мозга с подкреплявшими возбуждениями объясняет, почему при сохранности гипоталамических пейсмекеров повреждение других структур мозга не влияет критическим образом на поведение животных, т. к. при этом мотивационное возбуждение распространяется на сохранные отделы мозга. На самом деле это лишь видимость объяснения, апеллирующая к общему пониманию распределенности голографического явления. Действительно же объяснение кроется в том, что каждая фиксация текущего опыта происходит в различных областях мозга, ответственных за восприятие в данных специфических условиях внешнего мира и внутреннего состояния организма, что было хорошо изучено Н. Бехтеревой. Поэтому повреждения каких либо областей лишает мозг всего лишь опыта поведения в тех конкретных специфических условиях, что практически невозможно обнаружить. Все это открывает обнадеживающие перспективы компенсации поврежденных функций мозга.
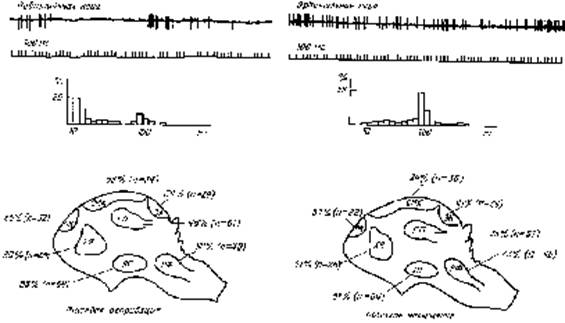
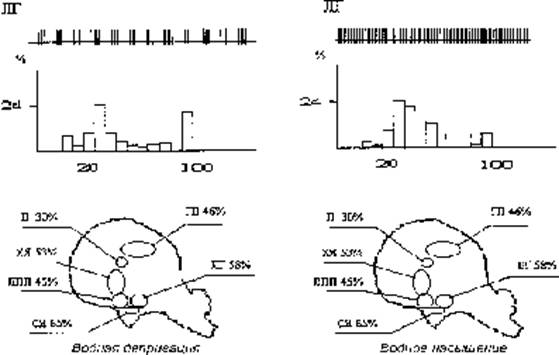
Рис. 2. Распределение нейронов с доминирующими межимпульсными интервалами по структурам мозга у кроликов при наличии мотивации голода и жажды и удовлетворении соответствующих потребностей. ЛГ – латеральный гипоталамус, ГП – дорзальный гиппокамп, ХЯ – хвостатое ядро, П – латеральное ядро перегородки, ЛПП – медиальное преоптическое поле перегородки, СЯ – супраоптическое ядро гипоталамуса.
Следует, однако, подчеркнуть, что голографический принцип построения функций мозга существенно отличается от физической голографии. Это признание обнадеживает:)
Физическая голограмма – “замороженный отпечаток действительности”. Голограммы мозга динамичны, они в зависимости от параметров подкрепляющих воздействий и различных исходных состояний организма все время изменяются. В живых организмах целесообразно говорить о динамической голографии функций мозга.
Взаимодействие доминирующих мотиваций с соответствующим подкреплением на структурах мозга включает эмоциональный компонент. Его происхождение связано с торсионным еще одно модное слово! механизмом деятельности функциональных систем. При формировании потребности, т.е. отклонении результата деятельности той или иной функциональной системы, под влиянием сигнализации о потребности, формируется отрицательная эмоция. При удовлетворении потребности, когда элементы функциональной системы работают в сторону возвращения результата к оптимальному для жизнедеятельности уровню, формируется положительная эмоция. Эмоции выступают, таким образом, в роли информационного пеленга состояний адаптивных результатов деятельности различных функциональных систем организма, и в частности, – гомеостатического уровня.
Структуры мозга, на которых осуществляется взаимодействие мотивационных и подкрепляющих возбуждений можно рассматривать в качестве своеобразного голографического информационного экрана. На этих структурах все время осуществляются интерференционные взаимодействия колебательных волн, вызванных мотивационными и подкрепляющими воздействиями (Судаков К.В., 2000). Вот и дошли до абсурда. В самом интересном месте не приводится даже малейшего обоснования "волнообразности" сигналом мозга, которые ни в коей мере таковыми не являются (в отличие от суперпозиции этих сигналов, воспринимаемых аппаратурой как энцефлографические ритмы). Однако, несмотря на вольную и популисткую интерпретацию, многие эксперименты и выводы автора очень интересны.
В качестве информационного экрана в каждой функциональной системе выступает аппарат акцептора результата действия. Именно, на структурах акцептора результата действия запечатляются свойства параметров подкрепляющих воздействий. Видимо предполагается, что информациюс этого экрана считывает нечто, вроде гомункулуса, с которым боролся Ф.Крик, упоминая в книге Мысли о мозге. Здесь, все же лучше не говорить популистски об "информационном" экране, а лучше просто сказать о профиле возбуждения.
Акцептор результата действия представляет в каждой функциональной системе разветвленную по структурам мозга архитектонику (Рис. 3). Его организация связана с распространением сформированных исходной потребностью мотивационных возбуждений на исполнительные пирамидные нейроны мозга. Одновременно с распространением этих возбуждений по пирамидному тракту на эффекторные органы, копии этих команд по коллатералям пирамидного тракта адресуются к вставочным нейронам различных областей мозга, в которых это возбуждение, благодаря замкнутым цепям, может длительно сохраняться. Именно к этим нейронам поступают потоки возбуждений (обратная афферентация) при действии на организм различных параметров подкрепляющих воздействий. При этом каждый параметр подкрепления запечатляется на специальной группе вставочных нейронов. В своей совокупности эти вставочные нейроны составляют целостный “образ” подкрепления – аппарат акцептора результата действия – отпечатки действительности. Мы полагаем, что указанные процессы лежат в основе динамических стереотипов высшей нервной деятельности, установленных И.П.Павловым.
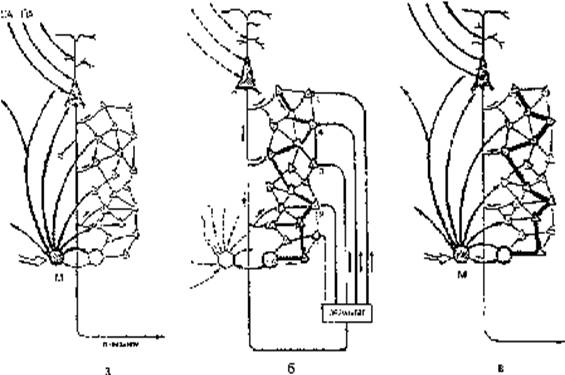
Рис. 3. Запечатление параметров подкрепления на структурах мозга, вовлеченных в доминирующую мотивацию и их последующее извлечение доминирующей мотивацией. а – распространение обусловленных доминирующей мотивацией (М) эффекторных возбуждений пирамидного тракта через его коллатерали на вставочные нейроны различных областей мозга, составляющие акцептор результатов действия; б – запечатление параметров подкрепления на структурах акцептора результата действия; в – последующее опережающее возбуждение доминирующей мотивацией структур акцептора результата действия.
Обогащение акцептора результата действия происходит всю жизнь субъекта. На ранних стадиях онтогенеза запечатляются признаки родителей и тех факторов внешней среды, которые удовлетворяют ведущие потребности новорожденных – явление импринтинга, открытое К.Лоренцем (Lorenz, 1965).
Основу запечатления составляют механизмы экспрессии ранних генов и, в частности, c-fos и c-jun (Анохин К.В., 1997). На фоне усиленной пищевой мотивации при исследовании новорожденными с помощью выраженной ориентировочно-исследовательской реакции окружающих предметов, у них наблюдается выраженная активация во многих структурах мозга гена c-fos. После достижения результата – удовлетворения исходной потребности – экспрессия гена c-fos снижается (Анохин К.В. с соавт., 2000).
При сканировании мозга с помощью позитронно-эмиссионного томографа обнаружено, что при наличии мотивации жажды у испытуемых в структурах поясной извилины, гиппокампа, таламуса, амигдалы и мозжечка наблюдается усиленный мозговой кровоток. После приема жидкости кровоток в этих областях мозга снижается (Denton D. et al., 1999).
Указанные процессы, по-видимому, составляют основу процессов запечатления параметров подкрепления на структурах акцептора результата действия. Интимная природа этих процессов все еще остается далеко не изученной, что требует проведения в этом направлении дальнейших научных исследований.
В формировании акцептора результатов действия в каждой функциональной системе ведущая роль принадлежит подкреплению (Судаков К.В., 1996).
Как показали опыты (Судаков C.K., 1987), молекулярная организация подкрепления включает процессы синтеза белка. Введение ингибиторов синтеза белка не подавляет у необученных животных ориентировочно-исследовательскую и оборонительную реакции при электрическом раздражении центров “голода” и “страха” соответственно латерального и вентромедиального гипоталамуса. Однако если вслед за электрическим раздражением указанных центров животные обучаются достигать полезный приспособительный результат, т. е. получать пищу при раздражении “центра голода” латерального гипоталамуса или избегать помещения, в котором им раздражали “центр страха” вентромедиального гипоталамуса, те же блокаторы синтеза белка при их введении в боковые желудочки мозга начинали эффективно подавлять указанные результативные реакции. Приведенные опыты свидетельствуют о том, что под влиянием подкреплений в структурах мозга формируются молекулярные энграммы подкрепления, блокируемые ингибиторами синтеза белка.
В проведенных нами опытах обнаружено, что пищевая мотивация у обученных кроликов, заблокированная ингибитором синтеза белка – циклогексимидом восстанавливалась при введении животным в боковые желудочки мозга пентагастрина, оборонительная реакция – при введении брадикинина, а реакция самораздражения – АКТГ4–10 (Судаков К.В., 1995).
Специальные опыты (Судаков С.К., 1987) показали, что указанные процессы формирования энграмм памяти разыгрываются на структурах акцептора результата действия.
Циклогексимид при его микроионофоретическом подведении блокировал у кроликов реакции вставочных нейронов сенсомоторной коры на антидромное раздражение пирамидного тракта.
Введение животным антигастриновых иммуноглобулинов также блокировало пищевые реакции у кроликов при раздражении “пищевого центра” латерального гипоталамуса (Судаков С.К., 1986). Все это также указывает на то, что в реализации доминирующей мотивации в соответствующее поведение принимают участие специальные белковые молекулы пентагастрина.
В извлечении сформированных подкреплением молекулярных энграмм акцептора результатов действия ведущая роль принадлежит доминирующим мотивациям. При возникновении соответствующей потребности, доминирующие мотивации опережающе извлекают сформированные ранее образы подкрепления. Этот механизм и направляет субъектов к цели – удовлетворению исходной доминирующей потребности.
Доминирующая мотивация настраивает мозг на более обостренное восприятие лучше сказать - на болееспецифичное, т.к. эти "мотивации" задают общий контекстдейст восприятию вительности и создает своеобразную “канву” возбужденных нейронов мозга, на которой подкрепляющие возбуждения “пишут” в каждом случае специфические энграммы, которые при очередном возникновении соответствующей потребности извлекаются доминирующей мотивацией. При блокаде молекулярных энграмм подкрепления мотивационное возбуждение, распространяющееся на структуры мозга, не способно сформировать соответствующее поведение.
Как показали опыты (Котов А.В. и Толпыго С.М., 1978), активация энграмм подкрепления также наблюдается под воздействием ранее выработанных условных раздражителей.
В процессе индивидуальной жизни происходит обогащение динамических стереотипов – соответствующих акцепторов результата действия.
Основой построения акцептора результата действия являются эмоциональные состояния. Они создают субъективный образ внутренних потребностей и их удовлетворения. Они же определяют эмоциональное сознание, строящееся на основе постоянной оценки потребностей и их удовлетворения.
Отпечатки действительности на структурах акцептора результатов действия динамичны и зависят от параметров подкрепления и способов удовлетворения потребностей.
Отпечатки действительности подобно “клубку шерсти” наматываются на ранее созданные стереотипы, таким образом, настоящая действительность неразрывно связывается с опытом прошлого.
Можно полностью согласиться с В.Гёте, который писал: “Все, что происходит с нами, оставляет тот или иной след в нашей жизни. Все участвует в создании нас такими, какие мы есть”.
Указанные процессы постоянного формирования на структурах мозга динамических стереотипов и извлечения из памяти “отпечатков действительности” и определяют, в конечном счете, интеллект личности.
С другой стороны, информационные процессы, разыгрывающиеся на структурах мозга, формируют субъективный опыт личности.
В архитектонику интеллекта включаются все стадии системной организации психической деятельности: доминирующая мотивация, обстановочная афферентация, память, принятие решения, предвидение потребных результатов (акцептор результатов действия), эфферентный синтез и, наконец, – оценка достигнутых параметров подкрепления с помощью акцептора результата действия. Однако при этом ведущую роль играет запечатление параметров результатов, удовлетворяющих исходные потребности организма на структурах акцептора результата действия, активированных доминирующими мотивациями.
Запечатление и воспроизведение доминирующей мотивацией отпечатков действительности – энграмм памяти в структурах акцептора результата действия – происходит на основе информационных эквивалентов действительности.
В 1969 г. П.К.Анохин писал: “…В теории информации существует представление о точности передачи информации о каком-либо объекте независимо от перекодирования. Я назвал бы эти этапы передачи информации информационным эквивалентом объекта. Это значит, что процесс информации, в каком бы звене передачи мы его ни уловили, принципиально содержит в себе все то, что составляет наиболее характерные черты исходного объекта, однако эти признаки могут быть представлены в разных кодах” (Анохин П.К., 1969).
Информационную основу интеллекта составляет эмоциональное сознание (Райков В.Л., 1998) – постоянная оценка разнообразных потребностей организма и их удовлетворения с помощью специфических эмоциональных переживаний.
В формировании субъективных образов объективной действительности можно выделить эндогенные и экзогенные стереотипы. Эндогенные стереотипы создаются на основе эмоциональных ощущений при возникновении и удовлетворении разнообразных внутренних (метаболических) потребностей организма. Экзогенные стереотипы формируются под воздействием факторов внешней среды, определяя сенсорное сознание. Кроме того, в общей архитектонике “отпечатков действительности”, наряду с сенсорными “отпечатками”, имеют место моторные стереотипы, связанные с трафаретными движениями, определяющими удовлетворение различных потребностей организма (Асмаян Н.В. и Богомолова Е.М., 1976). Всякому известно, как устойчивы заученные действия. Они проявляются нередко и вхолостую, когда по различным причинам изменяется ранее привычная обстановка. Этот элемент интеллекта можно рассматривать как “моторное сознание”.
Сформированные на эндогенной и экзогенной основах динамические стереотипы, характеризующиеся специфическими изменениями конвергентных и химических свойств нейронов мозга, выступают в роли голографических экранов мозга, своеобразного “фильтра” внешних воздействий.
У человека, в отличие от животных, субъективное восприятие действительности через динамические стереотипы определяется не только эмоциональными ощущениями, но и вербальным, языковым знанием. В этом проявляются первая и вторая сигнальные системы, предложенные И.П.Павловым.
Субъективная оценка на основе опыта внутренних состояний окружающей действительности определяет, в конечном счете, эмоциональное и вербальное сознание человека. Сознание выступает как процесс сопоставления информации, поступающей в мозг из внутренней и внешней среды с ранее накопленным информационным опытом – “отпечатками действительности”.
“…Конкретные механизмы рождения субъективного в сознании, – писал в 1969 году П.К.Анохин, – не поддаются пока аналитическому описанию, однако их точная информационная связь с исходными параметрами объективного мира не может быть подвергнута сомнению”.
Представления об информационных эквивалентах действительности позволяют уже реально приступить к изучению внутренней идеальной стороны – сознания человека.
Литература
Анохин К.В. Молекулярно-генетические предпосылки системогенеза поведенческих актов // Теория системогенеза / Под ред. К.В.Судакова. – М.: Горизонт, 1997. – С. 215–276.
Анохин К.В., Рябинин А.Э., Судаков К.В. Экспрессия гена c-fos в мозге у мышей в динамике выработки навыков оборонительного поведения // Журнал высш. нервн. деят. им. И.П.Павлова. – 2000.
Анохин П.К. Психическая форма отражения действительности // Ленинская теория отражения и современность / София: Науки и изкуство, 1969. – С. 109–139.
Анохин П.К. Философские аспекты теории функциональной системы // Избранные труды. – М.: Наука, 1978. – 400 с.
Анохин П.К. Кибернетика функциональных систем // Избранные труды. – М.: Медицина, 1998. – 400 с.
Асмаян Н.В., Богомолова Е.М. О возможных механизмах процесса принятия решения на примере условного инструментального поведения // Проблемы принятия решения / М.: Наука, 1976. – С. 171–177.
Громова Е.А. Эмоциональная память и ее механизмы. – М.: Наука, 1980. – 180 с.
Журавлев Б.В. Системный анализ активности нейронов мозга при пищедобывательном поведении у животных // Нейроны в поведении: системные аспекты / М., 1986. – С. 170–179.
Зилов В.Г. Интегративные свойства доминирующей мотивации на стадии афферентного синтеза // Вестник АМН СССР. – 1985 – № 2. – С. 93–96.
Котов А.В., Толпыго С.М. О роли пищевой мотивации в механизмах активации памяти у животных // Разгрузочно-диетическая терапия психических и соматических заболеваний с первично психическими нарушениями и ее физиологические механизмы / М., 1978. – C. 125–127.
Леонтьев А.Н. Деятельность. Сознание. Личность. – М.: Политиздат, 1976. – 304 с.
Павлов И.П. Рефлекс цели // Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных / М.: Медгиз, 1951. – С. 197–201.
Райков В.Л. Биоэволюция и совершенствование человека. Гипноз, Сознание, Творчество, Искусство. – М.: Изд. дом “Грааль”, 1998. – 648 с.
Сеченов И.М. Избранные философские и психологические произведения. – Госполитиздат, 1947. – С. 362.
Системокванты физиологических процессов / К.В.Судаков, Г.Ц.Агаян, Ю.Е.Вагин, С.М.Толпыго, Е.А.Умрюхин; Под ред. К.В.Судакова. – М.: Международный гуманитарный фонд Арменоведения им. акад. Ц.П.Агаяна, 1997. – 152 с.
Судаков К.В. Олигопептиды в системных механизмах поведения // Физиологически активные пептиды. – Пущино, 1988. – С. 68–79.
Судаков К.В. Пейсмекер доминирующей мотивации // Физиол. журнал им.
И.М.Сеченова. – 1992. – Т. 78. – № 12. – С. 1–11.
Судаков К.В. Гипоталамические пейсмекеры в механизмах формирования алкогольного влечения: роль олигопептидов // Вестник новых медицинских технологий. – Тула, 1994. – Т. 1. – № 2. – С. 53–58.
Судаков К.В. Мотивация и подкрепление в системных механизмах поведения: динамические энграммы подкрепления // Журнал высш. нервн. деят. им. И.П.Павлова. – 1995. – Т. 45. – В. 4. – С. 627–637.
Судаков К.В. Системный процесс подкрепления // Журнал высш. нервн. деят. им. И.П.Павлова. – 1996. – Т. 46. – В. 4. – С. 643–655.
Судаков К.В. Голографический принцип системной организации процессов жизнедеятельности // Успехи физиол. наук. – 1997. – Т. 28. – № 4. – С. 3–32.
Судаков К.В. Биологические мотивации в системной организации функций мозга // Журнал невролог. и психиатрии им. С.С.Корсакова. – 1998. – Т. 98. – № 2 – С. 53–58.
Судаков К.В. Системная организация функций головного мозга: определяющая роль акцептора результатов действия // Журнал неврол. и психиатрии им. С.С.Корсакова. – 1998. – Т. 98. – № 4. – С. 13–19.
Судаков К.В. Индивидуальная устойчивость к эмоциональному стрессу. – М.: Горизонт, 1998. – 267 с.
Судаков К.В. Системная организация функций человека: Теоретические аспекты // Успехи физиол. наук. – 2000. – Т. 31. – № 1. – С. 1–17.
Судаков К.В. Информационный феномен жизнедеятельности. – М., 2000.
Судаков К.В., Бадиков В.И., Зилов В.Г., Федянина Н.Г. Исследование физиологических свойств мотивациогенных центров гипоталамуса в условиях действия олигопептидов // Проблемы физиологии гипоталамуса. – Киев, 1987. – Вып. 21. – С. 3–8.
Судаков С.К. Нарушение пищевого поведения кроликов при внутрижелудочковом введении антигастриновых иммуноглобулинов // Журнал высш. нервн. деят. им. И.П.Павлова. – 1986. – Т. 36. – № 2. – С. 391.
Судаков С.К. Возможная роль окситоцина и вазопрессина как факторов доминирования мотиваций // Ж. высш. нервн. деят. – 1987. – Т. 37. – Вып. I. – С. 160–164.
Судаков С.К. Молекулярные механизмы памяти в системной организации поведенческих актов // Функциональные системы организма. – М.: Медицина, 1987. – С. 166–178.
Судаков С.К. Молекулярные механизмы включения пирамидных нейронов сенсомоторной коры мозга в организацию пищевого поведения кроликов // Нейрофизиология. – 1987. – Т. 19. – № 5. – С. 601–606.
Урываев Ю.В. Системный анализ функций лобной коры: Автореф. дисс. д-ра мед. наук. – М., 1978. – 31 с.
Ухтомский А.А. Доминанта как фактор поведения // Собр. соч., 1925. – T. I. – С. 293.
Ясперс К. Общая психопатология: Пер. с нем. – М.: Практика, 1997.
Denton D. et al. Neuroimaging of genesis and satiation of thirst and an interoceptor-driven theory of origins of primary consciousness // Proc. Natl. Acad. Sci. USA. – 1999. – V. 96. – P. 5304–5309.
Lorenz К. Evolution and modification of behavoir. – Chicago-Lond.: Univ. Chicago Press, 1965 – 121 p.
Robbins T.W., Everit B.J., Muir J.L., Harrison A. IBRO News, 1992. – V. 20. – N 3. – P. 7.
Summary
System Basis of the Intellect
K.V.Sudakov
P.K.Anokhin Institute of Normal Physiology, Russian Academy of Medical Sciences, Moscow
The paper presents data testifying to new aspects in understanding brain functions on the basis of the theory of functional systems, postulated by P.K.Anokhin.
Unlike a widespread conductive neurotransmission paradigm of brain function organization, cortical-subcortical interactions stipulated by the activity of the multiple functional systems of the organism are regarded.
It is shown that in every given moment of time the brain activity is being built by a dominant motivation. Holographic mechanism of interaction between motivation and reinforcement on brain structures is of a special consideration.
A statement, that dominant motivations act as a structure-functional “groundwork”, where reinforcing stimulation forms “the imprinting of the reality” i.e. dynamic stereotypes, is postulated.
A hypothesis about the role of the apparatus of the acceptor of action result as a holographic screen of functional systems is also under consideration in the paper. Enrichment of the acceptor of action result during an individual’s life moulds a subject’s intellect when evaluation of various parameters of the reality by the acceptor of action result forms a subject’s consciousness.