Свойства генетического кода - след его возникновения.
Относится к разделу Молекулярная биология
К серии статей, описывающих происхождение ГК, можно относиться как к расследованию событий, о которых у нас осталось очень немало следов. Однако для понимания этих статей необходимо немного приложить усилий для вникания в молекулярные механизмы синтеза белка. Данная статья является вступительной для серии автопубликаций, посвященных возникновению генетического кода, и с неё лучше всего начинать знакомство с этой темой.
Обычно генетический код (ГК) определяют как способ (правило) кодирования белка на первичной структуре ДНК или РНК. В литературе чаще всего пишут, что это - однозначное соответствие последовательности из трёх нуклеотидов в гене одной аминокислоте в синтезируемом белке или месту окончания синтеза белка. Однако в таком определении есть две ошибки. При этом подразумеваются 20, так называемых канонических аминокислот, которые входят в состав белков всех без исключения живых организмов. Эти аминокислоты являются мономерами белка. Ошибки следующие:
1) Канонических аминокислот не 20, а только 19. Аминокислотой мы можем называть вещество, которое одновременно содержит аминогруппу -NH2 и карбоксильную группу - COOH. Дело в том, что мономер белка - пролин - аминокислотой не является, поскольку в нём вместо аминогруппы присутствует иминогруппа, поэтому пролин правильней называть иминокислотой. Однако в дальнейшем во всех статьях, посвящённых ГК, для удобства я буду писать о 20 аминокислотах, подразумевая указанный ньюанс. Структуры аминокислоты приведены на рис. 1.
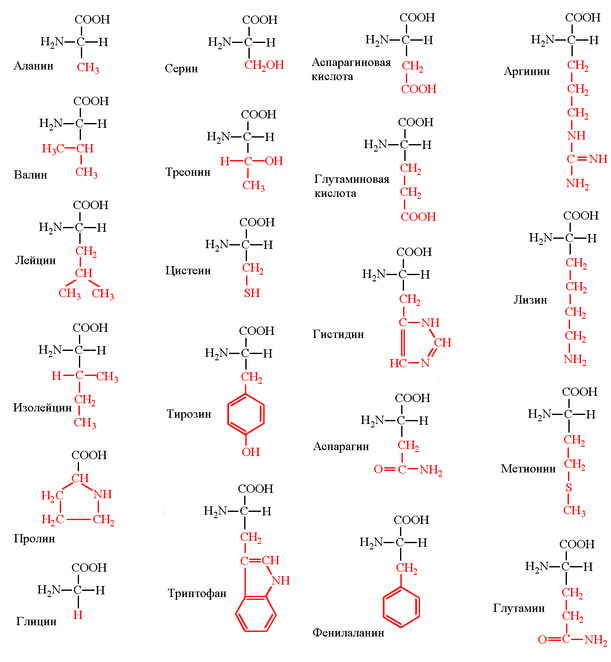
Рис. 1. Структуры канонических аминокислот. Аминокислоты имеют константные части, обозначенные на рисунке чёрным цветом, и вариабельные (или радикалы), обозначенные красным.
2) Соответствие аминокислот кодонам не всегда является однозначным. О нарушении случаев однозначности см. ниже.
Возникновение ГК означает возникновение кодируемого синтеза белка. Это событие является одним из ключевых для эволюционного формирования первых живых организмов.
Структура ГК представлена в круговой форме на рис. 2.
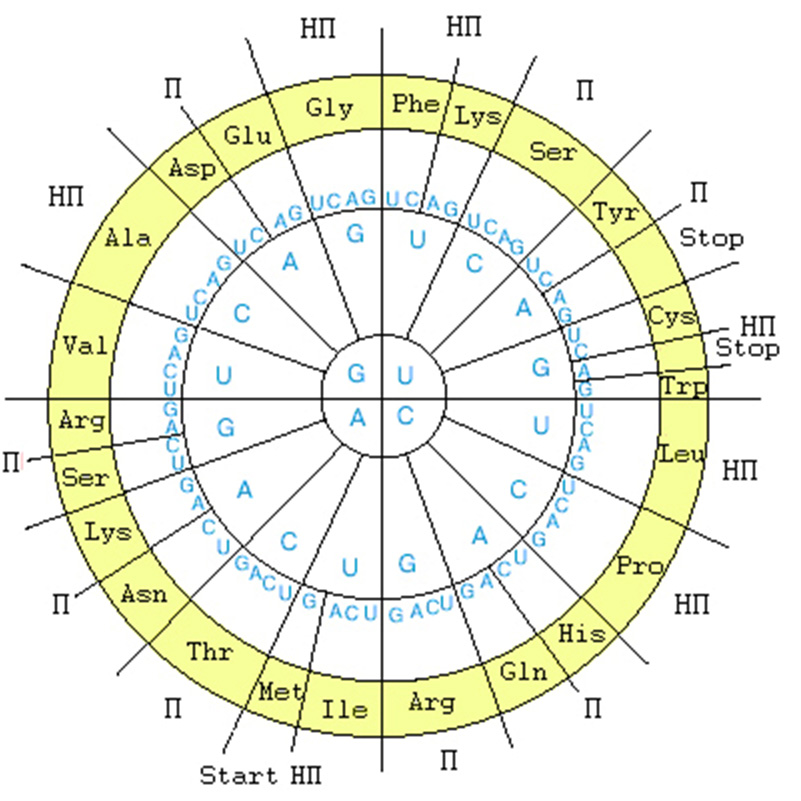
Рис. 2. Генетический код в круговой форме. Внутренний круг - первая буква кодона, второй круг - вторая буква кодона, третий круг - третья буква кодона, четвертый круг - обозначения аминокислот в трехбуквенном сокращении; П – полярные аминокислоты, НП – неполярные аминокислоты. Для наглядности симметрии важен избранный порядок символов U-C-A-G.
Итак, приступим к описанию основных свойств ГК.
1. Триплетность. Каждая аминокислота кодируется последовательностью из трёх нуклеотидов.
2. Наличие межгенных знаков препинания. К межгенным знакам препинания относятся последовательности нуклеиновой кислоты, на которых трансляция начинается или заканчивается.
Трансляция может начаться не с любого кодона, а только со строго определённого – стартового. К стартовому кодону относится триплет AUG, с которого начинается трансляция. В этом случае этот триплет кодирует или метионин, или другую аминокислоту – формилметионин (у прокариот), который может включаться только в начале синтеза белка. В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляцию (так называется синтез белка на рибосоме).
3. Компактность, или отсутствие внутригенных знаков препинания. Внутри гена каждый нуклеотид входит в состав значащего кодона.
4. Неперекрываемость. Кодоны не перекрываются друг с другом, каждый имеет своё упорядочённое множестов нуклеотидов, которое не перекрывается с аналогичными множествами соседних кодонов.
5. Вырожденность. Обратное соответствие в направлении аминокислота-кодон неоднозначно. Это свойство называется вырожденностью. Серия – это множество кодонов, кодирующих одну аминокислоту, другими словами, это группа эквивалентных кодонов. Представим себе кодон в виде XYZ. Если XY определяет “смысл” (т.е. аминокислоту), то кодон называется сильным. Если же для определения смысла кодона нужен определенный Z, то такой кодон называется слабым.
Вырожденность кода тесно связана с неоднозначностью спаривания кодон-антикодон (под антикодоном подразумевается последовательность из трёх нуклеотидов на тРНК, которая может комплементарно спариваться с кодоном на матричной РНК (см. более подробно об этом две статьи: Молекулярные механизмы обеспечения вырожденности кода и Правило Лагерквиста. Физико-химическое обоснование симметрий и соотношений Румера). Один антикодон на тРНК может узнавать отодного до трёх кодонов на мРНК.
6. Однозначность. Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляции.
Известно три исключения.
Первое. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин. В начале гена формилметионин кодируется как обычным метиониновым кодоном AUG, так и ещё валиновым кодоном GUG или лейциновым UUG, которые внутри гена кодируют валин и лейцин, соответственно.
Во многих белках формилметионин отщепляется, либо удаляется формильная группа, в результате чего формилметионин превращается в обычный метионин.
Второе. В 1986 году сразу несколько групп исследователей обнаружили, что на мРНК терминирующий кодон UGA может кодировать селеноцистеин (см. рис. 3) при условии, что за ним следует особая последовательность нуклеотидов.
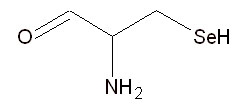
Рис. 3. Структура 21-й аминокислоты - селеноцистеина.
У E. coli (это латинское название кишечной палочки) селеноцистеил-тРНК в процессе трансляции распознает в мРНК кодон UGA, но лишь в определенном контексте: для узнавания UGA-кодона как осмысленного важна последовательность длиной в 45 нуклеотидов, расположенная вслед за UGA-кодоном.
Рассмотренный пример показывает, что при необходимости живой организм может изменять смысл стандартного генетического кода. В этом случае генетическая информация, заключенная в генах, кодируется более сложным образом. Смысл кодона определяется в контексте с определенной протяженной последовательностью нуклеотидов и при участии нескольких высокоспецифических белковых факторов. Важно, что селеноцистеиновая тРНКобнаружена в представителях всех трёх ветвей жизни (архей, эубактерий и эукариот), что указывает на древность происхождения селеноцистеинового синтеза, и возможно на присутствие его ещё в последнем универсальном общем предке (о нём речь пойдёт в других статьях). Скорей всего селеноцистеин встречается у всех без исключения живых организмов. Но в каждом отдельном организме селеноцистеин встречается не более, чем в паред есятков белков. Он входит в состав активных центров ферментов, в ряде гомологов которых на аналогичной позиции может функционировать обычный цистеин.
До недавнего времени считалось, что кодон UGA может считываться либо как селеноцистеин, либо кактерминальный, но недавно было показано, что у инфузории Euplotes кодон UGA кодирует или цистеин, илиселеноцистеин. См. ""
Третье исключение. У некоторых прокариот (5 видов архей и одной эубактерии - в Википедии информация сильно устарела) встречается особая кислота – пирролизин (рис. 4). Она кодируется триплетом UAG, который в каноническом коде служит терминатором трансляции. Предполагается, что в этом случае, подобно случаю с кодированием селеноцистеина, считывание UAG как пирролизинового кодона происходит благодаря особой структуре на мРНК. Пирролизиновая тРНК содержит антикодон CTA и аминоацилируется АРСазой 2-го класса (про классификацию АРСаз см. статью "Кодазы помогают понять, как возник генетический код").
UAG в качестве стоп-кодона используется редко, а если и используется, то часто за ним следует другой стоп-кодон.
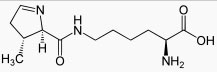
Рис. 4. Структура 22-й аминокислоты пирролизина.
7. Универсальность. После того, как в середине 60-х годов прошлого века расшифровка ГК была завершена, долгое время считалось, что код одинаков во всех организмах, что указывает на единство происхождения всего живого на Земле.
Попробуем понять, почему ГК универсален. Дело в том, что если бы в организме изменилось хотя бы одно правило кодирования, то это привело бы к тому, что изменилась структура значительной части белков. Такое изменение было бы слишком кардинальным и поэтому практически всегда летальное, так как изменение смысла только одного кодона может затронуть в среднем 1/64 часть всех аминокислотных последовательностей.
Отсюда следует одна очень важная мысль - ГК почти не менялся со времени своего формирования более 3,5 млрд. лет назад. А, значит, его структура несёт в себе след его возникновения , и анализ этой структуры может помочь понять, как именно мог возникнуть ГК.
В действительности ГК может несколько отличаться у бактерий, митохондрий, ядерный код некоторых инфузорий и дрожжей. Cейчас насчитывают не менее 17 генетических кодов, отличающихся от канонического на 1-5 кодонов Суммарно во всех известных вариантах отклонений от универсального ГК используются 18 различных замен смысла кодона. Больше всего отклонений от стандартного кода известно у митохондрий - 10. Примечательно, что митохондрии позвоночных, плоских червей, иглокожих, кодируются разными кодами, а плесневых грибков, простейших и кишечнополостных - одним.
Эволюционная близость видов - отнюдь не гарант того, чтобы у них были сходные ГК. Генетические коды могут различаться даже у разных видов микоплазм (одни виды имеют канонический код, а другие - отличающиеся). Аналогичная ситуация наблюдается и для дрожжей.
Важно отметить, что митохондрии - потомки симбиотических организмов, которые приспособились жить внутри клеток. Они имеют сильно редуцированный геном, часть генов переселилась в ядро клетки. Поэтому изменения ГК в них становятся уже не столь кардинальными.
Обнаруженные позднее исключения представляют особый интерес с точки зрения эволюции, поскольку могу помочь пролить свет на механизмы эволюции кода.
Таблица 1.
Митохондриальные коды у различных организмов.
Кодон | Универсальный код | Митохондриальные коды | |||
Позвоночные | Беспозвоночные | Дрожжи | Растения | ||
UGA | STOP | Trp | Trp | Trp | STOP |
AUA | Ile | Met | Met | Met | Ile |
CUA | Leu | Leu | Leu | Thr | Leu |
AGA | Arg | STOP | Ser | Arg | Arg |
AGG | Arg | STOP | Ser | Arg | Arg |
Три механизма смены аминокислоты, кодируемой кодом.
Первый – когда какой-то кодон не используется (или почти не используется) каким-то организмом в силу неравномерности встречаемости каких-то нуклеотидов (GC-состав), или комбинаций нуклеотидов. В результате такой кодон может вовсе исчезнуть из употребления (например, благодаря потере соответствующей тРНК), а в дальнейшем может использоваться для кодирования другой аминокислоты без нанесения существенного ущерба организму. Этот механизм возможно отвечает за появление некоторых диалектов кодов у митохондрий.
Второй – превращение стоп-кодона в смысловой. В этом случае часть у части транслируемых белков могут появиться дополнения. Однако ситуацию частично спасает то, что многие гены часто заканчиваются не одним, а двумя стоп-кодонами, поскольку возможны ошибки трансляции, при которых стоп-кодоны считываются как аминокислоты.
Третий – возможно неоднозначное считывание определённых кодонов, как это имееют место у некоторых грибов.
8. Связность. Группы эквивалентных кодонов (то есть кодонов, кодирующих одну и ту же аминокислоту) называются сериями. ГК содержит 21 серию, включая стоп-кодоны. В дальнейшем для определенности любая группа кодонов будет называться связной, если от каждого кодона этой группы можно перейти ко всем другим кодонам этой же группы путем последовательных замен нуклеотидов. Из 21 серии связны 18. 2 серии содержат по одному кодону, и лишь 1 серия для аминокислоты серин является несвязной и распадается на 2 две связные подсерии.
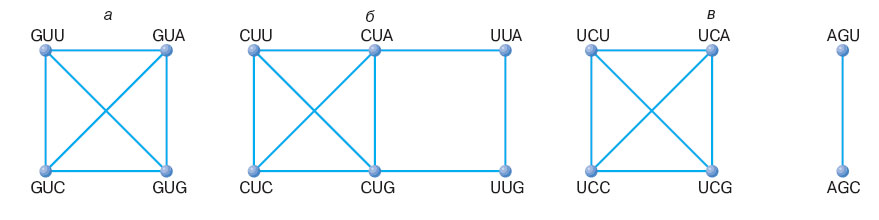
Рис. 5. Графы связности для некоторых кодовых серий. а - связная серия валина; б - связная серия лейцина; серия серина несвязная, распадается на две связных подсерии. Рисунок взят из статьи В.А. Ратнера "".
Свойство связности можно объяснить тем, что в период формирования ГК захватывал новые кодоны, которые минимально отличались от уже используемых.
9. Регулярность свойств аминокислот по корням триплетов. Все аминокислоты, кодируемые триплетами скорнем U, являются неполярными, не крайних свойств и размеров, имеюталифатические радикалы. Все триплеты с корнем C имеют сильные основы, ааминокислоты, кодируемые ими, имеют относительно малые размеры. Все триплеты с корнем A имеют слабые основы, кодируют полярные аминокислоты не малых размеров. Кодоны с корнем G характеризуются крайними и аномальнными вариантами аминокислот и серий. Они кодируют самую маленькую аминокислоту (глицин), самую длинную и плоскую (триптофан), самую длинную и «корявую» (аргинин), самую реактивную (цистеин), образует аномальную подсерию для серина.
10. Блочность. Универсальный ГК является «блоковым» кодом. Это означает, что аминокислоты со сходными физико-химическими свойствами, кодируются кодонами, отличающимися друг от друга одним основанием. Блочность кода хорошо видна на следующем рисунке.
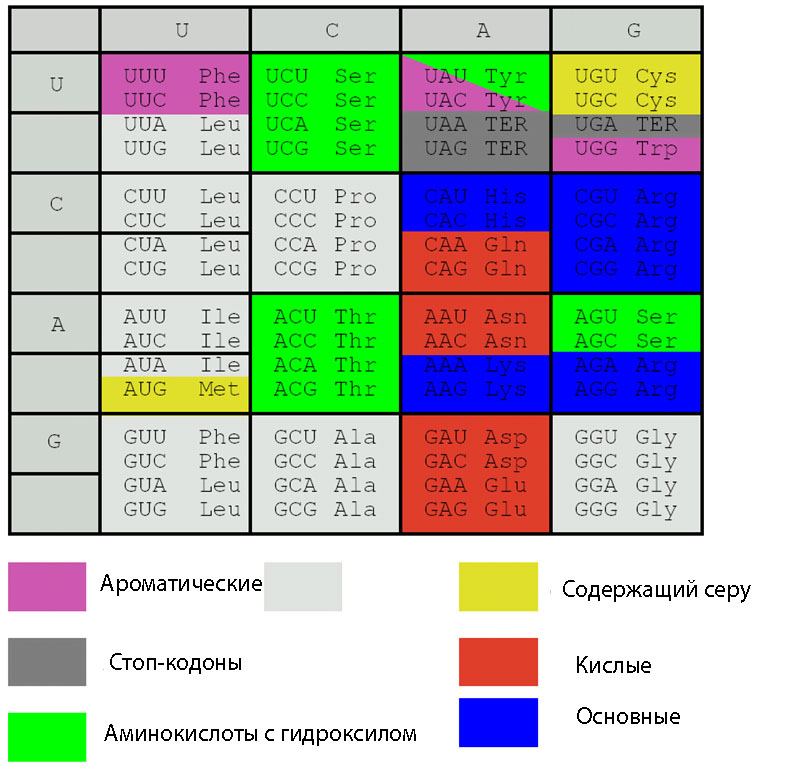
Рис. 6. Блочная структура ГК. Белым цветом обозначены аминокислоты с алкильной группой.

Рис. 7. Цветовое представление физико-химических свойств аминокислот, основанное на значениях, описанных в кн книге Стайерса "Биохимия" [Berg et al. 2002]. Слева - гидрофобность. Справа - способность к формированию альфа-спирали в белке. Красный, жёлтый и голубой цвета обозначают аминокислоты с большой, средней и малой гидрофобностью (слева) или соответствующей степенью способности к формированию альфа-спирали (справа).
Свойство блочности и регулярности также можно объяснить тем, что в период формирования ГК захватывал новые кодоны, которые минимально отличались от уже используемых.
Кодоны с одинаковыми первыми основаниями (приставками кодонов) кодируют аминокислоты с близкими путями биосинтеза [Tailor F.J., Coates D. 1989]. Кодоны аминокислот, принадлежащих к шикиматному, пируватному, аспартатному и глутаматному семействам, имеют в качестве приставок U, G, A и C, соответственно. О путях древнего биосинтеза аминокислот и его связи со свойствами современного кода см. "Древний дублетный генетический код был предопределён путями синтеза аминокислот". На основе этих данных некоторые исследователи делают вывод о том, что на формирование кода большое влияние оказали биосинтетические взаимоотношения между аминокислотами [Wong J.T. 1975, Amirnovin R. 1997]. Однако сходство биосинтетических путей вовсе не означает сходство физико-химических свойств [Di Giulio M. 1997b].
11. Помехоустойчивость. В самом общем виде помехоустойчивость ГК означает, что при случайных точковых мутациях и ошибках трансляции не очень сильно меняются физико-химические свойства аминокислот.
Замена одного нуклеотида в триплете в большинстве случаев или не приводит к замене кодируемой аминокислоты, или приводит к замене на аминокислоту с той же полярностью.
Один из механизмов, обеспечивающих помехоустойчивость ГК – его вырожденность. Средняя вырожденность равна - число кодируемых сигналов/общее число кодонов, где к кодируемым сигналам относятся 20 аминокислот и знак терминации трансляции. Усредненная вырожденность для всех аминокислот и знака терминации составляет три кодона на кодируемый сигнал.
Для того, чтобы количественно оценить помехоустойчивость, введём два понятия. Мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными. Мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
Каждый триплет допускает 9 однократных замен. Всего кодирующих аминокислоты триплетов 61. Поэтому количество возможных замен нуклеотидов для всех кодонов –
61 x 9 = 549. Из них:
23 замены нуклеотидов приводят к появлению стоп-кодонов.
134 замены не меняют кодируемую аминокислоту.
230 замен не меняют класс кодируемой аминокислоты.
162 замены приводят к смене класса аминокислоты, т.е. являются радикальными.
Из 183 замен 3-его нуклеотида, 7 приводят к появлению терминаторов трансляции, а 176 - консервативны.
Из 183 замен 1-ого нуклеотида, 9 приводят к появлению терминаторов, 114 - консервативны и 60 - радикальны.
Из 183 замен 2-го нуклеотида, 7 приводят к появлению терминаторов, 74 -консервативны, 102 - радикальны.
На основе этих расчётов получим количественную оценку помехоустойчивости кода, как отношение числа консервативных замен к числу радикальных замен. Оно равно 364/162=2.25
При реальной оценке вклада вырожденности в помехоустойчивость необходимо учитывать частоту встречаемости аминокислот в белках, которая варьирует в разных видах.
В чем причина помехоустойчивости кода? Большинство исследователей считают, что это свойство является следствием селекции альтернативных ГК [Freeland S.J., Hurst L.D. 1998].
Стивен Фриленд и Лоренс Херст генерировали случайные такие коды и выясняли, что только один из ста альтернативных кодов обладает не меньшей помехоустойчивостью по сравнению с универсальным ГК.
Еще более интересный факт обнаружился, когда эти исследователи ввели дополнительное ограничение, с тем чтобы учесть реально существующие тенденции в характере мутирования ДНК и появлении ошибок при трансляции. При таких условиях лучше канонического кода оказался ТОЛЬКО ОДИН КОД ИЗ МИЛЛИОНА ВОЗМОЖНЫХ .
Столь беспрецедентную жизнестойкость генетического кода проще всего объяснить тем, что он сформировался в результате естественного отбора. Возможно когда-то в биологическом мире существовало множество кодов, каждый со своей чувствительностью к ошибкам. Организм, лучше справлявшийся с ними, имел больше шансов выжить, и канонический код просто победил в борьбе за существование. Это предположение кажется вполне реальным - ведь мы знаем, что альтернативные коды действительно существуют. Подробнее о помехоустойчивости см. (С.Фриленд, Л. Херст "Закодированная эволюция".//В мире науки. – 2004, №7).
В заключение, предлагаю посчитать число возможных генетических кодов, которые можно сгенерировать для 20 канонических аминокислот. Почему-то это число нигде мне не попадалось. Итак, нам необходимо, чтобы в генерируемых ГК были обязательно 20 аминокислот и стоп-сигнал, кодируемые ХОТЯ БЫ ОДНИМ КОДОНОМ.
Мысленно будем нумеровать кодоны в каком-то порядке. Рассуждать будем следующим образом. Если у нас имеется ровно 21 кодон, то тогда каждая аминокислота и стоп-сигнал будут занимать ровно по одному кодону. В этом случае возможных ГК будет 21!
Если будет 22 кодона, то появляется лишний кодон, который может иметь один из любых 21 смыслов, причём этот кодон может располагаться на любом из 22 мест, тогда как остальные кодоны имеют ровно по одному разному смыслу, как и для случая 21 кодонов. Тогда получим число комбинаций 21!х(21х22).
Если кодонов будет 23, то рассуждая аналогично, получим, что 21 кодон имеют ровно по одному разных смыслов (21! вариантов), а два кодона - по 21 разных смысла (212 смыслов при ФИКСИРОВАННОМ положении этих кодонов). Число различных положений для этих двух кодонов будет 23х22. Общее число вариантов ГК для 23 кодонов - 21!х212х23х22
Если кодонов будет 24 - то число ГК будет равно 21!х213х24х23х22,...
....................................................................................................................
Если кодонов будет 64, то число возможных ГК будет 21!х2143х64!/21! = 2143х64! ~ 9.1х10145
А в колоде-то всего 4 масти. 
ЛИТЕРАТУРА
Berg J.M., Tymoczko J.L., & Stryer L. Biochemistry 5th ed. New York: W. H. Freeman and Company. 2002.
Статьи, связанные с темой:
Биохимическая (коэволюционная) модель эволюции генетического кода
Древний дублетный генетический код был предопределён путями синтеза аминокислот
Кодазы помогают понять, как возник генетический код
Молекулярные механизмы обеспечения вырожденности кода
Список публикаций >>
Тест: А не зомбируют ли меня? Тест: Определение веса ненаучности
| Обнаружен организм с крупнейшим геномом Новокаледонский вид вилочного папоротника Tmesipteris oblanceolata, произрастающий в Новой Каледонии, имеет геном размером 160,45 гигапары, что более чем в 50 раз превышает размер генома человека. | Тематическая статья: Формирование субъективного опыта в онтогенезе |
Рецензия: Работы А.Иваницкого, комментарии | Топик ТК: Свойства осознанного внимания и циклы |