Автотрофное термофильное происхождение жизни.
Относится к разделу Молекулярная биология
"До сегодняшнего дня на Земле активную роль играют солнечная энергия, энергия гравитационных взаимодействий и энергия радиоактивного превращения элементов внутри Земли. Конвенктивные массы переносят тепло с помощью этих трёх форм энергий от самого ядра нашей планеты до верхних слоёв атмосферы, в конечном счёте обеспечивая согласованное взаимодействие химического напряжения и питательных компонентов жизни. Без конвенкции земной мантии не было бы ни тектоники, ни вулканов, ни гор, ни горячих источников, а значит, - ни одной новой поверхности, которая обеспечивала бы образование химического потенциала, способного двигать древние метаболические процессы."
[Russell M.J., Hall, A.J., Mellersh A.R. 2003]
В различных моделях абиогенного происхождения жизни на Земле по разному рассматривается происхождение синтеза ключевых веществ, необходимых для синтеза информационных молекул и компонентов их метаболизма. В данном вопросе на современном уровне знаний не может быть окончательной точки зрения. Долгое время наиболее популярной была гетеротрофная модель происхождения первых живых систем, идущая ещё с работ А.И. Опарина и Дж. Холдейна. Согласно гетеротрофной модели источником углерода для первых живых систем были уже готовые абиогенно синтезируемые органические молекулы. Автотрофные же процессы сформировались позднее после истощения первичного бульона.
Радикально другой взгляд просматривается в автотрофной модели происхождения жизни, согласно которой фиксация углерода из углекислого газа была возможна уже в самых первых живых системах, которые "взяв под контроль" какие-то геохимические реакции с помощью наследственно предопределённого синтеза новых ферментов, сформировали первичный автотрофный метаболизм, то есть управляемую фиксацию углерода из углекислого газа.
Первые во многом можно даже сказать ещё немногословные, если не сказать робкие высказывания относительно роли гидротермальных источников, были озвучены в очень короткой заметке в журнале "Nature" в 1988 М.Расселом с соавт. [Russell M.J. et al. 1988].
Его одна из первых статей по этой теме [Wächtershäuser G. 1988с] начинается с отрицания. Он отрицает:
1. Первичный бульон, то есть часто используемое представление о наличии некой смеси сложных органических веществ в водоёмах
древней Земли. Должны были отсутствовать начальные строительные блоки для сложных органических молекул. Даже такие, как аминокислоты.
2. Связанное с этим бульоном формирование первых гетеротрофных систем.
3. Присутствие других «миров» - прообразов современных живых систем, таких, как РНК-мир или мир кристаллической глины.
Фактически Г. Вехтерхойзер предложил модель возникновения двумерных живых систем, поскольку они должны были существовать на поверхности минералов. Согласно его модели идею первичного бульона более уместно заменить моделью "примитивной пиццы". По этой теории жизнь на земле возникла на поверхности железно-серных минералов, то есть сульфидов и пиритов, которые и сегодня образуются посредством геологических процессов, а на молодой земле должны были встречаться гораздо чаще. Жизнь согласно этой модели должна была возникнуть в форме мономолекулярного слоя, состоящего из взаимодействующих анионных метаболитов, электростатически связанных с положительно заряженными поверхностями минералов в горячей воде [Wächtershäuser G. 1988a]:
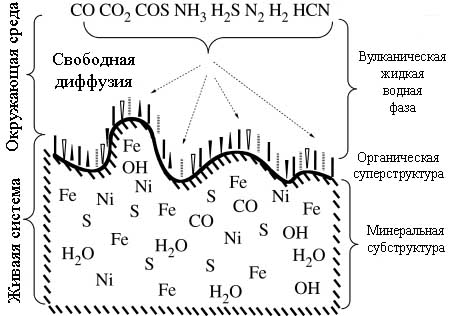
Рис. 1. Двумерная организация первых живых систем. Рисунок взят из [Wächtershäuser G. 2006] c небольшими изменениями.
Ключевой идеей его модели является предположение о том, что положительно заряженные двухвалентные катионы железа и отрицательно заряженные анионы серы адсорбируются на поверхности минералов и реагируют с образованием пирита:
Fe2+ + 2H2S → FeS2 + 4H+ + 2e-.
Данная реакция являетя существенно экзергонической (см. внизу в словаре свободная энергия (энергия Гиббса)), что обеспечивает принципиальную возможность для протекания изначально эндергонических реакций.
По своей сути эта модель предполагает, что первый биокатализ был гетерогенным. Термин "гетерогенный катализ" означает, что, во-первых, химические реакции, ставшие прообразом первого метаболизма, катализировались на поверхности раздела фаз, а, во-вторых, что протеканию химических реакций предшествовал процесс адсорбции веществ на поверхности раздела сред.
Дальнейшее разлвитие каталитической активности в отсутствие полимерных РНКовых и белковых катализаторов происходило в направлении от гетерогенного катализа к так называемым гомогенному и микрогетерогенному катализу.
Этот первичный катализ привёл к синтезу малых органических молекул, вроде лимонной, яблочной кислоты, различных тиокислот и тиоэфиров (об этом подробнее ниже).
Эти вновь появившиеся молекулы - эффективные (с точки зрения катализа) лиганды, которые могли связываться с ионами переходных металлов, в результате чего катализ мог осуществляться уже в растворе и быть гомогенным (без раздела фаз) или же микрогетерогенным (в котором, во-первых, участвовали уже не настоящие растворы, а коллоидные, а, во-вторых, подобно гетерогенному катализу, протеканию реакции предшествовал процесс адсорбции малых органических молекул на поверхности уже растворённых железо-серных кластеров. Подробнее об этом см. Минеральные корни современных ферментов
Первые автокаталитические метаболические циклы на поверхности минералов должны были раскрутить новые метаболические сети и образование первых полимерных наследственных молекул.
В данной модели образование комплексных биомолекул связано с постоянным надежным источником энергии.
Одним из ключевых свойств ЖС является наличие в ней так называемых сопряжённых химических реакций. Суть явления сопряжения сводится к тому, что одни экзергонические реакции высвобождают энергию для протекания других реакций. В предложенной Вехтерхойзером модели сопряжение реакций заключается в том, что энергия поступает из восстановления частично окисленных железно-серных минералов, например пирита (FeS2), водородом и поставляет достаточно энергии для эндотермического синтеза мономерных структурных элементов биомолекул и их полимеризации. В качестве источника углерода рассматривается СО2 или СО - оба этих газа выделяются в больших количествах при извержениях вулканов.
Экспериментально обнаруженные химические реакции, протекающие с участием металлосерных соединений.
Взаимодействие FeS с H2S в присутствии CO2 в анаэробных условиях даёт не только водород, но и разнообразные органические серусодержащие вещества, что и послужило основой для выдвижение Г.Вехтерхойзером модели железо-серного метаболизма. При этом экспериментально удалось подтвердить связывание CO2 с образованием серусодержащих органических веществ, таких как СS2, различные тиолы и метилсульфиды [Heinen W., Lauwers A.M. 1996].
К важнейшим химическим реакциям, протекающим вблизи горячих источников являются перечисленные ниже.
(1) Образование пиритов и молекулярного водорода из сульфидов и СО2 [Taylor P et al. 1979]:
FeS + H2S = FeS2 + H2
(2) Окисление СО до СО2 [Wächtershäuser G. 1988c; Seewald J. S. et al. 2006] происходит как в присутствии различных катализаторов, так и без них в вулканических флюидах:
СО + Н2О = СО2 + 2Н+ + 2е;
(3) Окисление железных соединений, например, гидроксида [ Huber C., Wächtershäuser G. 2003]:
2Fe(OH)2 = 2FeO(OH) + 2H+ +2e
(4) Молекулярный водород составляет существенную часть как современных, так и древних гидротермальных потоков [ Kelley D.S. et al. 2001], а в древнем океане Гадея его содержание должно быть даже выше [Russell M.J., Hall A.J. 1997]. Окисление H2 [Wächtershäuser G. 1988a,c]:
H2 = 2H+ + 2e
FeS/H2S-фиксация азота, предсказанная сначала Вехтерхойзером теоретически [Wächtershäuser G. 1988c], а затем продемонстрированная экспериментально [Dörr M et al. 2003a,b]:
N2 + 3FeS + 4H2S = 3FeS2 + 2NH3;
(5) Катализируемое никелем образование СOS из H2S [Huber C. Wächtershäuser G. 1997]:
CO + H2S = COS + 2H+ + 2e
(6) Образование метилмеркаптана из CO2 и FeS/H2S [Heinen W., Lauwers A.M. 1996]:
CO2 + 3FeS + 4H2S = CH3SH + 3FeS2 + 2H2O.
(7) Образование метилмеркаптана из CO и H2S на NiS [Huber C. Wächtershäuser G. 1997]:
СO + H2S + 4e + 4H+ = CH3SH + H2O
(8) Образование активированной тиоуксусной кислоты (CH3-CO-SH) из CH3SH и СО как эволюционного предшественника тиоэфиров уксусной кислоты было предсказано теоретически [Wächtershäuser G. 1990], а затем продемонстрировано экспериментально при 1000С [Huber C., Wächtershäuser G. 1997]:
2СH3SH + CO = CH3-CO-S-CH3 + H2S
Показано, что эта реакция может катализироваться (Fe,Ni)S, NiS, Ni(OH)2 и CoS.
Полученное вещество и его биосинтез в некотором роде напоминают современный ацетил-CoA. Подобно ацетил-СоА данное вещество может рассматриваться и как гипотетический универсальный строительный материал, и как универсальный источник энергии.
Немаловажным также является и образование трёхуглеродных органических соединений.
(9) Возможно также, что формирование некоторых компонентов более современного аналога ВЦТК было обусловлено участием фотосинтетических процессов.
Как выяснилось недавно, пять реакций ВЦТК могут протекать в отсутствие белковых катализаторов и катализироваться минералами. Так, сравнительно недавно было показано, что 5 реакций ВЦТК могут катализироваться на поверхности минерала сфалерита (ZnS) фотохимически, то есть с поглощением энергии света [Xiang V. Zhang, Scot T. 2006]. При этом катализируются обе реакции укороченного варианта восстановительного цикла лимонной кислоты, в которой происходит фиксация углекислого газа.
Из данных, связанных с анализом роли пиритов железа, цинка и других металлов ещё предстоит выкристаллизоваться более детальной экспериментальной модели происхождения древнего ВЦТК.
(10) Образование пирувата фиксацией CO было сначала предсказано теоретически [Huber C. Wächtershäuser G. 1997], а затем продемонстрировано экспериментально при давлении 2000 бар при температуре 2500С c участием FeS/RSH [ Cody G.D. 2000]:
3CO + 4H+ + 4e = CH2-CO-CO-OH
Данная реакция образования пирувата может рассматриваться как предковый прообраз современного синтеза пирувата при восстановительном карбоксилировании ацетил-СоА.
(11) Образование α-аминокислот из α-кетокислот было вначале предсказано теоретически [Wächtershäuser G. 1990], а затем экспериментально продемонстрировано при 1000С [Hennet R.J.C. et al. 1992; Hafenbradl D. et al. 1995; Huber C.,Wächtershäuser G. 2003]:
R-CO-COOH + 2Fe2+ + NH3 + 2H+ = R-CH(NH2)-COOH + 2Fe3+ + H2O
Реакция происходит с участием FeS и (или) Fe(OH)2 как катализаторами и источник электронов (электроны передаются при окислении железа от степени окисления +2 до +3. За окислением железа следует образование FeS2 в присутствии H2S или FeOOH, Fe2O3, Fe3O4 и др. в отсутствие сероводорода.
Внимание! Возможный реликтовый след! Глутамат-синтаза в современных организмах для катализирования восстановительного аминирования также использует FeS-кластеры для транспорта электронов.
(12) Активация аминокислот и пептидный цикл. Образование пептидной связи из активированных аминокислот было открыто при добавлении в реакционную смесь аминокислот для образования тиоэфиров уксусной кислоты. В присутствии (Fe,Ni)S, CO и H2S (или CH3SH) at 100°C реакция протекает лучше в щелочных условиях при использовании Mg(OH)2 как буфера. COS может рассматриваться как активирующий интермедиат, так как он вместо СО также может давать пептиды [Huber C.,Wächtershäuser G. 1998]. Была также обнаружена активация аминокислот с помощью СOS при комнатной температуре в отсутствие (Fe,Ni)S [Lenan L. et al. 2004].
Однако в пептидах N-терминальные аминокислоты в присутствии СО преобразуются в которое затем превращается в мочевину. В результате возможен гипотетический пептидный цикл превращения аминокислот, который (что очень важно для модели перехода в олигорибонуклеиновый мир!) потенциально способен давать предшественники для синтеза пуриновых нуклеотидов. Схема простейшего гипотетического глицин-глицинового цикла приведена на рис. 2. На первом шаге глицин активируется через его N-карбоксиангидрид реакцией с COS. В этой реакции используется энергия окислительно-восстановительной реакции (3). Далее аминогруппа другой молекулы глицина реагирует с аминоацил-N-карбоксиангидридом, вновь давая дипептид глицин-глицин. Последний, в свою очередь, опять же реагирует с СOS(CO/H2S), генерируя как аналог N-карбоксиангидрида.
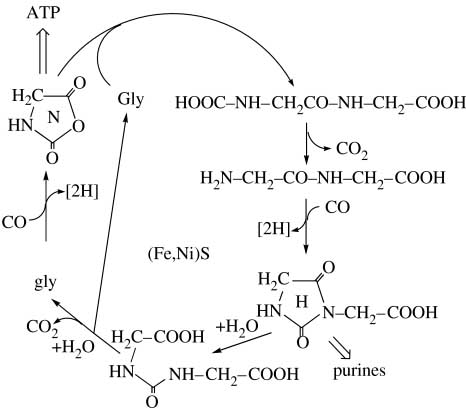
Рис. 2. Дипептидный цикл глицина. Рисунок взят из [Wächtershäuser G. 2006].
Важно, что конкурируя с образованием гидантоина, свободные аминогруппы дипептида могут реагировать с другой молекулой N-карбоксиангидрида, образуя трипептид.
Таким образом, в рамках данной гипотетической модели уже подразумевается наличие олигопептидов, которые могли существенно расширить функциональные возможности эволюционирующих первых автокаталитических систем, являющихся уже первыми ЖС. Г.Вехтерхойзер предполагает, что аминогруппа могла связываться с металлами-лигандами, которая предотвращает превращение N-терминальной аминокислоты в гидантоин [Wächtershäuser G. 2006], что требует дальнейшего экспериментального подтверждения. Сами олигопептиды (или металлоолигопептиды) можно рассматривать как компоненты конформационного генома (о том, чем отличается конформационный геном от полимерного - см. "Что было первым: метаболизм или ген? Могла ли жизнь начаться как циклическая химическая реакция неполимерных молекул?"), способный к дарвиновской эволюции.
Кроме того, пептидный цикл может рассматриваться как каталитический цикл, родстенный СО-дегидрогеназе. Источником энергии для данного цикла являются молекулы СО (их окислительно-восстановительный потенциал).
(13) Реакции фосфоририрования. Вулканические газы содержат летучий P4O10, который гидролизуется через полифосфатные и пирофосфатные интермедиаты [Yamagata Y. et al. 1991]. Пирофосфаты уже несколько десятилетий рассматриваются как предковые фосфорилирующие агенты [Baltscheffsky H. 1971].
Возможны также дополнительные важнейшие реакции, которые могут протекать при олигорибонуклеиновом контроле (в рамках рассматриваемой в излагаемом цикле статей древней стадии олигорибонуклеинового мира). Немаловажным для рассмотрения последующей эволюции кодируемого пептидного синтеза оказывается тот факт, что синтезируемый в рассматриваемых условиях COS может обеспечивать реакцию аминокислоты с AMP, с образованием аминокислоты-АМР, а также с фосфатом, с образованием аминоацил-фосфата, который впоследствии реагирует с другими фосфатами и пирофосфатами [Leman L. et al. 2006].
Таким образом, в настоящее время можно говорить не только о возможности фиксации углерода в горячих железо-серных источниках, но и о возможности запуска автокаталитических реакций, обладающих конформационным геномом. Одни из потенциальных кандидатов на роль последних - архаичный прообраз ВЦТК, пептидный цикл. Ниже в статье будут рассмотрены и другие возможные кандидаты на роль архаичных циклов. Актуальной остаётся пролема нахождения путей синтеза рибонуклеотидов, как главных кандидатов на роль рибоолигонуклеиновго мира - предшественника РНК-пептидного мира. Рассмотренные реакции образуют по-видимому верхушку айсберга, который ещё предстоит экспериментально изучить более глубоко.
Рассмотренные реакции могли объединяться в метаболические пути, грубая гипотетическая схема которых изображена на рис. 3:
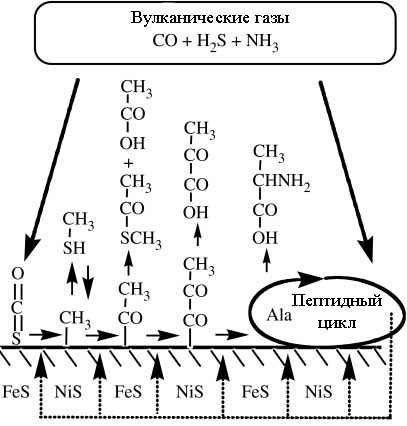
Рис. 3. Интегрированная гипотетическая сеть реакций с автокаталитической обратной связью (пунктирные стрелочки). Рисунок взят из [Wächtershäuser G. 2006] с небольшими изменениями.
К вопросу о компартментализации ЖС.
Принцип компартментализации живых клеток означает, что биохимические процессы в клетке локализованы в определённых отсеках, покрытых оболочкой, чаще из бислоя липидов. Для эволюции первых ЖС изоляция отдельных геномов (полимерных или конформационных) была необходимым условием для дарвиновского отбора. Поэтому возникает очень актуальный вопрос - как у первых ЖС, с недостаточно развитыми путями биосинтеза могли возникнуть компартменты?
Одно из решений этого вопроса в рамках гидротермального возникновения жизни было предложено М. Расселом и В. Мартином [Russell M.J., Martin W. 2004]. Щелочные флюиды древнейших гидротерм выносили водород, сульфиды разных металлов и аммиак, а также ионы таких металлов, как железо и никель, сульфиды которых в изобилии осаждались вокруг источников. Микроскопические поры в минеральной корке таких сульфидов, как грейгит, предоставляли полуизолированное пространство для синтеза как относительно простых органических молекул, так и более сложные пептиды (например, в пептидном цикле). Эти пептиды покрывали внутреннюю поверхность пор, и в связи с описываемой моделью гидротермального происхождения жизни их можно рассматривать как предковые аналоги современных ферментов, содержащих металло-серные кластеры.
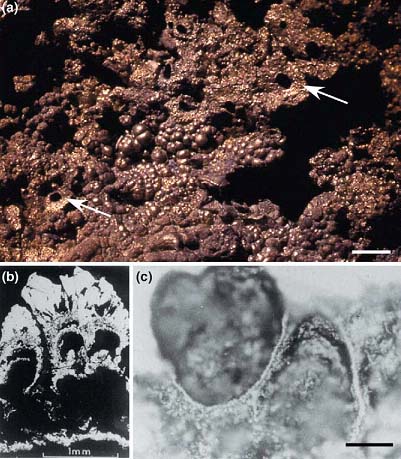
Рис. 4. Компартмены, формируемые железо-серными соединениями в горячих источниках.
(а) Внешний вид железо-сероных осадков гидротермального происхождения на дне моря (Тина, Ирландия, возраст около 350 млн. лет),. Поверхность содержит пузырьки (ботриоиды) и отверстия (показаны стрелочками), через которые выходили гидротермальные воды
(b,с) Микрофотографии ботриоидов, демонстрирующие металл-сульфидные компартменты.
Длина полоски для верхней фотографии соответствует 1 см, для фотографии (c) - 100 мкм.
Рисунок взят из [Russell M.J., Martin W. 2004].
Данные компартменты могли служить своеобразными химическими фабриками, в которых синтезировались органические вещества. Источником для синтеза для них служили выходящий из гидротермальных вод молекулярный водород, концентрация которого в гадейский период была даже более высокой, нежели в современную эпоху [Russell M.J., Hall A.J 1997], соприкасавшегося с CO2, концентрация которого также была существенной для гадейского океана [Nisbet E.G., Sleep, N.H. 2001]. Стенки этих компартментов кроме изоляции служили катализаторами химических реакций, обеспечивавших синтез органических веществ.
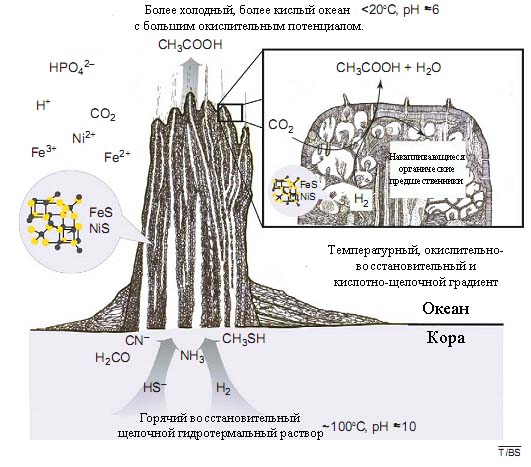
Рис. 5. Гидротермальный химический реактор, образованный на дне гадейских водоёмов щелочными горячими источниками (наподобие современных "белых курильщиков", см. объяснение далее в статье). В данных районах создавался сущетвенный температурный (от 110 до 200С), pH (от 10 до 6) и окислительно-восстановительный (с потенциалом от -600 мВ до +100 мВ) градиенты. Реактор имел форму горы, образованной карбонатами, глинами, оксидами, гидроксидами и сульфидами железа, никеля и других металлов. Образующиеся ионизированные и полярные органические вещества частично накапливались внутри "реакторов". Слабоионизированные и слабополярные органические вещества (такие, как уксусная кислота) выходили в океан.
Рисунок взят из [Russell M.J., Martin W. 2004].
Согласно данной модели первые ЖС представляли собой фактически каменные ячейки, в которых протекали циклические химические реакции, ставшие прообразом современного промежуточного метаболизма (цикл Кребса, ацетил-коэнзимА-путь, гидрокси-пропионатный путь и др.). Сами же минералы такие как грейгит, макинавит, виоларит и др. являлись прообразами активных центров современных ферментов и могли катализировать уже значительную часть древнего первого метаболизма (подробнее об этом см. в статье Минеральные корни современных ферментов). Такого рода предковый минеральный метаболизм стал прообразом того, что Эдвардс назвал универсальным предковым метаболическим комплексом (universal ancestral metabolic complex, UAMC).
Трудно представить, чтобы такого рода ячейки могли бы эффективно воспроизводиться. Поэтому, не смотря на важность данного этапа одним из ключевых этапов эволюции жизни должно было стать образование клеточной мембраны и клеточной стенки прокариот. Не случайно, что клеточная мембрана имеет независимое происхождение у архей и бактерий, что говорит об их достаточно позднем образовании.
По моему мнению клеточная стенка из вещества, наподобие пептидогликана должна была образоваться существенно раньше, об этом будет сказано в отдельной статье.
Пока лишь отмечу, что живые организмы покинули каменные ячейки, когда научились сами строить клеточную стенку (см. рис. 10).
Долгое время в качестве возможных кандидатов на синтез органики рассматривались так называемые "чёрные курильщики" (о них подробнее см. ниже). Однако из-за высокой температуры (около 3500С) количество и разнообразие синтезируемых в них органических веществ оказалось невысоким по сравнению с более холодными "белыми курильщиками" [Russell M.J., Hall A.J. 1997].
Гидротермальные воды гадейского периода (как и нынешние) содержат сероводород (H2S и HS-) в существенных концентрациях, который соприкасаясь с ионами as Fe2+ и Ni2+, являвшихся существенными компонентами гадейского океана [Russell M.J., Hall A.J. 1997]. Этот процесс обеспечивал выпадение осадков. Нынешний океан вместо Fe2+ содержит более окисленное железо Fe3+ из-за насыщения впоследствии поверхности Земли молекулярным кислородом.
Существенный кислотно-щелочной градиент (значение рH в гидротрмальных водах и океане различаетс на 4 единицы - см. рис. 8) в изолированных сульфидных кампартментах должен был обеспечить образование хемоосмотического протонного потенциала, который в современных организмах является связанное с мембраной универсальной химической валютой наряду с АТР (о трёх универсальных энергетических валютах см. статью В.П. Скулачёва "").
Наконец такого рода кампартментализация - это решение так называемой концентрационной проблемы возникновения жизни - возможности создания локально высоких концентраций органических веществ.
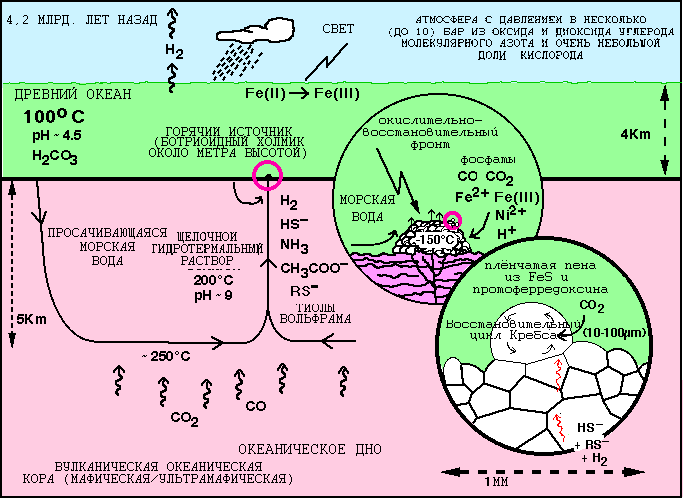
Рис. 6. Геохимические условия, в которых возникли первые биохимические циклы. Первые биохимические циклы возникли в плёнчатых ботриодиных (то есть имеющих форму виноградной кисти) образованиях, содержащих FeS, в которых железо в сульфидах принимало участие в переносе электрона как первый аналог ферредоксина, благодаря чему в этих мембранных образованиях образовывался водородный потенциал. В результате геохимической эволюции внутри ячеек возник прообраз восстановительного цикла Кребса.
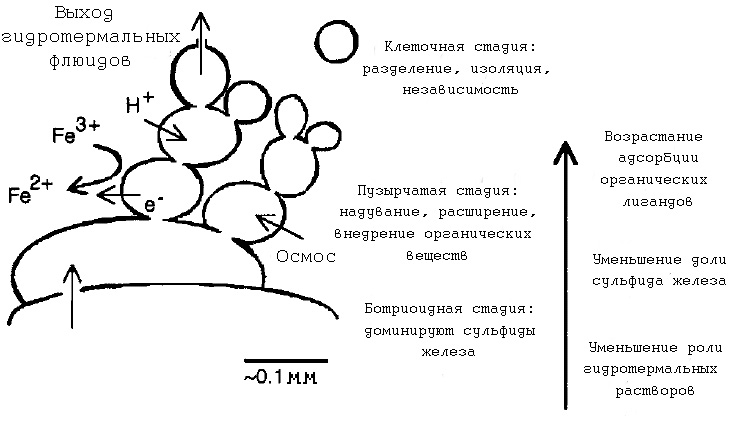
Рис. 7. Различные стадии эволюции компартментов. Рисунок взят из [Russell M.J. et al. 1993].
Гипертермофилы лежат в основании дерева жизни.
Все современные живые организмы на нашей планете делятся на три царства: Archaea, Bacteria, Eucarya. Анализ различных последовательностей белков и генов позволяет судить о вертвлении филогенетического дерева. Такого рода анализ выявляет одну существеннейшую особенностью, затрагивающую все три царства этого филогенетического древа: все имеющие глубокие корни ветви находятся между современными гипертермофилами (организмами, которые живут при 80-110°С и больше и обычно не развиваются при температуре ниже 60°С) [Nisbet E.G. Fowler C.M.R. 1996; Stetter, K. O. 1996].
Оказалось, что для гипертермофильных архей вольфрам является абсолютно необходимым элементом жизни, тогда как для мезофилов этот элемент может быть необязательным.
В свете рассмотренных моделей присутствие этогор элемента в археях представляет своего рода "ископаеую биохимию" и означает, что в гипотетических минеральных ячейках должен был присутствовать вольфрам, что подтверждается его нахождением в щелочных гидротермах в виде растворённых тиолов в форме WIVS32- (рис. 9).
Неожиданное разнообразие гидротермальных источников.
Курильщики - подводные образования вокруг выходов из ложа океана термальных вод. Впервые так называемые "чёрные курильщики", получившие своё название благодаря чёрым клубам растворов сульфидов металлов. Обнаружены в 1970-х гг. в Красном море, затем — в Тихом и Атлантическом океане. Эта группа гидротермальных источников является как в настоящее время, так и на начальных этапах формирования жизни по видимому является наиболее многочисленной. Черные курильщики» встречаются вдоль границ тектонических плит, где выходят на поверхность рудоносные горячие насыщенные газами компоненты магмы с температурой до 375 — 550 °C. Эта температура существенно ниже температуры магмы (около 8000С) из-за частичного испарения воды [Von Damm K.L. 2000]. Соприкасаясь с водой на выходе, вода остывает и сульфиды (в основном железа и марганца), находящиеся в ней, выпадают в осадок. Отложения сульфидов имеют форму конусов. Кроме того, в выходах термальных вод содержатся в значимых количествах золото, медь, цинк, свинец и другие металлы. Высота образований — до 100 м.
Такая высокая температура гидротермали обеспечивает закисливание выходящей воды до pH.
Mg2+ + 2H2O → Mg(OH)2 (силикат) + 2H+


Рис. 8. Чёрные курильщики. (Фото geo.uni-bremen.de).
Долгое время они рассматривались в качестве единственного аналога древних гидротермальный процессов. В них вода обладает кислотное реакцией (pH 1-3), которая обусловлена растворённой в ней соединений серы, дающих серную кислоту. Такого рода соединения рассматриваются как причина того, что более 4 млрд лет назад только что сформированный древний океан, содержащий предположительно во много раз меньше воды, был кислым, и кислотность эта поддерживалась также существенной концентрацией углекислотго газа в атмосфере. Важной особенностью чёрных курильщиков является выделение больших объемов CO2, H2S и ионов металлов.
Некоторые исследователи обращали внимание, что чёрные курильщики проблематично рассматривать как колыбели жизни из-за того, что высокие температуры в них не позволяют синтезировать сложные органические вещества [Bada J.L. 2004; Orgel L.E. 2008]. В качестве дополнителного аргумента выдвигалось также их относительно небольшое время жизни (несколько десятилетий) [ Kelley D.S. et al 2002].
Однако наряду с высокотемпературными чёрными курильщиками вулканического происхождения были также обнаружены чёрные курильщики невулканического происхождения, обнаруженные к западу от Марианского желоба. По-видимому, они являются выходами ювенильных вод (образованы в результате соединения выделившихся из магмы кислорода и водорода и дегидратации пород в зоне метаморфизма) или холодных вод, являющихся побочным продуктом образования серпентенита, идущего в толще пород.
В последние 20 лет стало ясно, что горячие подводные источники намного разнообразней, нежели предполагалось ранее. Обнаружение новых типов горячих источников можно назвать "последним крупным географическим открытием". В середине 80-х годов, были найдены также остатки древних щелочных источников [Fallick A.E., Ilich M., Russell M.J. 1991]. Наконец в 2000-м году в 15 км от срединного океанического хребта в Атлантическом океане в подводном горном массиве были описаны принципиально новые гидротермальные источники, которые в отличие от чёрных курильщиков не обязательно сосредоточены на границах тектонических плит.. Эти источники обладают щелочной реакцией и являются относительно "холодными": их температура может доходить до 200 °C, но обычно варьирует от 70 до 1500С [Kelley D.S. et al. 2001]. Они были обнаружены как белые конусы высотой до 60 м, рядом с которыми находятся образования более причудливой формы, а также многочисленные расщелины, заполненные белой породой. Эти термальные источники нового типа были названы «белыми курильщиками» или «Затерянным городом» (англ. Lost City). «Белые курильщики» сложены из карбонатных пород. Вода, изливающаяся из этих «курильщиков», нагревается за счет энергии, выделяющейся при реакции, протекающей между растворенными в морской воде веществами и оливином (веществом, составляющим значительную часть океанической коры). Этот процесс получил название серпентизация.
Важно, что эти гидротермальные системы не являются вулканическими. Во время этой реакции оливин превращается в другой минерал — серпентин; при этом выделяются метан, водород и избыточное тепло. Соединения серы и железа в них почти не присутствуют. Изливающаяся вода разогрета до 50 — 80 °C. В осадок выпадают такие минералы, как кальцит, арагонит (имеет подобно известняку и мрамору формулу CaCO3) и брукит. Соединения серы и железа почти не присутствуют (вместо сульфидов присутствуют небольшие количества сульфатов).Над "белыми башнями" не витает облака дыма. Поэтому эти источники распознаются только по световым бликам в местах выхода водяных струй из глубины.
Щелочную реакцию источникам придаёт карбонат кальция, который при выносе горячих минерализованных струй в холодные вóды океана, выпадает в осадок, напоминающий снег и образовывающие длинные наросты (десятки метров), напоминающие башни. Удивительным и необычайно важным оказался тот факт, что в этих источниках присутствует органика абиотического происхождения, включая метан и ацетаты, а также молекулярный водород [Proskurowski G.; et al.2008]. Присутствие метана потенциально может служить источником для синтеза других органических веществ.
Процесс серпентизации сопровождается выделением и других восстановителей помимо водорода, которых достаточно для фиксации молекулярного азота N2 и восстановление его до аммиака [Dörr M et al. 2003a,b], что потенциально даёт возможность синтез азотных интермедиатов, необходимых для биосинтеза аминокислот и нуклеотидов.
Важность именно щелочных гидротермальных источников как возможных колыбелей жизни была предсказана М. Расселом с соавт. почти за 10 лет до их открытия [Russel M.J. et al. 1993, 1994]. Этот новый тип гидротем важен не только своей щелочной реакцией, создающей градиент pH в древнем океане, но и также наличием тонких поровых структур с арагонитовыми стенками микроновых размеров.
Немаловажным фактом является также то, что белые курильщики должны были быть существенно более многочисленными на дне океана древней Земли и иметь время жизни на порядок большее, чем чёрные курильщики [Martin W., Russell M. 2003; Russell MJ, Arndt NT. 2005].
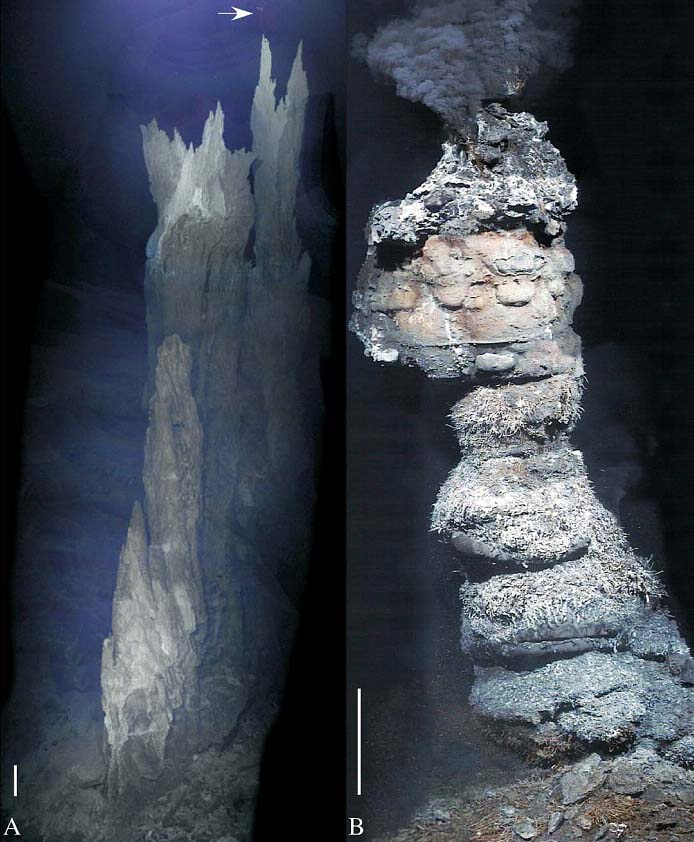
Рис. 9. Различия между карбонатной башней в "Затерянном городе" и (слева) и "трубой" чёрного курильщика (справа). Длина отрезка соответствует 1 м. Фотографии взяты из
И чёрные, и белые курильщики являются местами обитания экзотических экосистем, основу которых составляют хемоавтотрофы. Причина такой концентрации заключается не только в наличии эффективных доноров (H2) и акцепторов электронов (CO2), но и по-видимому в наличии абиогенно образованного органического углерода. Предполагаемая обобщённая схема фиксации углерода в белых курильщиках представлена на рис. 13.
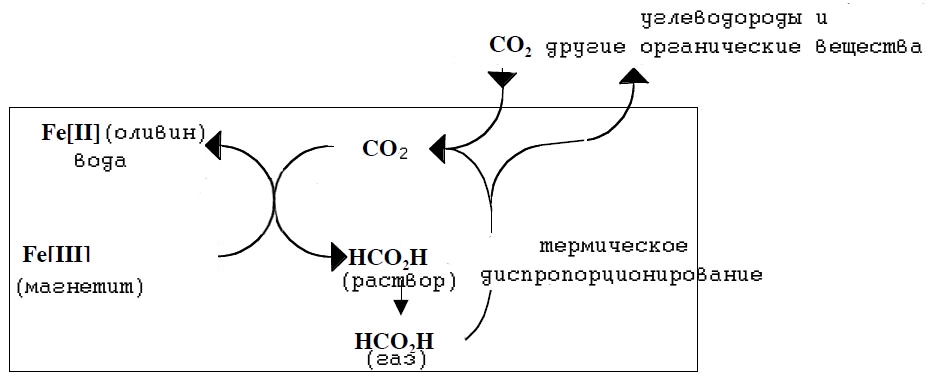
Рис. 10. Фиксация углерода в белых курильщиках, сопряжённая с процессом серпентизации (переходом магнетита в оливин).
Впервые факт фиксации углерода в присутствии оливина экспериментально подтвердил Т. МакКолом в 2000 году [McCollom T.M. 2000]. Он показал, что бикарбонат-ионы восстанавливаются до формата при 3000С и давлении 300 бар. Образующаяся муравьиная кислота может далее диспропорционировать с образованием углеводородов и диоксида углерода:
(3n+1) HCO2H → (2n+1) CO2 + CnH2n+2 + 2n H2O
Что говорят геологические данные?
Очень существенным фактом из геологии является наличие высокой концентрации диоксида углерода. В гадейскую эру (ранее 3,8 млрд. лет) его концентрация была в тысячу раз выше по сравнению с сегодняшней.
Отсутствие кислорода обеспечивало присутствие ионов железа, растворённых в океане.
Одной из особенностью связывания неорганического углерода является непропорциональное включение изотопов углерода. В результате этого процесса изотопный состав меняется в сторону уменьшения доли изотопа 13С. Поэтому изотопный анализ ископаемых пород позволяет оценить время происхождение автотрофного метаболизма. Фракционирование изотопов углерода 12С и 13С в древних осадочных породах показало, что автотрофный метаболизм возник примерно в то же время, когда Земля стала пригодной для существования жизни - около 3,8 млрд. лет назад [Schidlowski M. et al. 1983].
Справочные данные.
Альдольная конденсация - взаимодействие двух альдегидов, приводящее к возникновению альдегидоспирта.
Енольная форма - в карбонильных соединениях - альдегидах и кетонах - равновесная форма, в которой подвижный атом водорода от атома углерода переходит к кислороду:
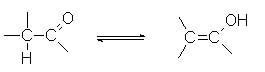
Оливи́н — породообразующий минерал, магнезиально-железистый силикат с формулой (Mg,Fe)2[SiO4] (железо двухвалентное). Один из самых распространённых на земле минералов.
Свободная энергия (энергия Гиббса) — физическая величина, выражаемая через энтальпию, температуру и энтропию системы G = H – T × S. Является термодинамическим потенциалом, то есть её изменение зависит только от начального и конечного состояния термодинамической системы. Её изменение позволяет нам судить, пойдёт ли реакция самопроизвольно, или нет. Если ΔG < 0, то данная реакция сможет протекать самопроизвольно, такие реакции называются экзергоническими. Если же ΔG > 0, то прямая реакция самопроизвольно идти не может (такие реакции носят название эндергонических), зато сможет идти обратная реакция. Нетрудно заметить, что у экзотермических реакций, в ходе которых выделяется тепло, ΔH < 0, следовательно, экзотермичность способствует самопроизвольному протеканию реакции. Увеличение энтропии в результате реакции также является фактором, уменьшающим ΔG, и, следовательно, благоприятствует ее протеканию.
Серпентизация — реакция ультрамафических (ультраосновных) пород, богатых оливином и пироксеном с водой, в результате которой образуется молекулярный водород и минералы из группы серпентина.
Серпентизация пироксена (приводит к увеличению величины pH до 10 и более [Macleod G. et al. 1994]):
12Ca0.25Mg1.5Fe0.25Si2O6 + 16H2O → 6Mg3Si2O5(OH)4 + 12SiO2 + Fe3O4 + 3Ca2+ + 6OH- + H2↑
Серпентизация оливина (приводит к образованию метана):
6Mg2SiO4 + 12Fe2SiO4 + 14H2O + CO2 → 8Fe3O4 + 4Mg3Si2O5(OH)4 + 10SiO2 + CH4↑ + 4H2↑
Ультрамафические (ультраосновные) горные породы — породы с относительно малым содержанием кремнезёма, SiO2, (< 45 мас.%) и обогащённые магнием и железом. Сосредоточены в верхней мантии, в которой представлены преимущественно перидотиты (главным образом, два минерала - оливин (Mg,Fe)2[SiO4] и пироксен (Ca,Na)(Fe,Mg,Al)(Si,Al)2O6). любое твёрдое вещество, поведение которого при деформации может быть описано как поведение жидкостей. Все реальные твёрдые тела под действием тангенциальных нагрузок ведут себя как жидкости, если время действия внешней силы, вызывающей в теле касательные напряжения, больше времени релаксации. В геологических процесса, длительность которых измеряется сотнями, тысячами и тем более миллионами лет, в качестве флюида могут выступать не только газы, водные растворы, неффть, илы, магма, но и глины, соли, гипсы, ангидриды, известняки и другие "твёрдые" тела.
Флюид — любое вещество, поведение которого при деформации может быть описано как поведение жидкостей. Все реальные тела, какими бы твёрдыми они не казались, под действием длительных тангенциальных нагрузок ведут себя как жидкости, если время действия внешней силы, вызывающей в теле касательные напряжения, больше времени релаксации. В геологических процессах, длительность которых нередко измеряется миллионами лет, в качестве флюида могут выступать не только газы, водные растворы, нефть, илы, магма, но и глины, соли, гипсы, ангидриды, известняки и другие "твёрдые" вещества.
Литература.
На в pdf-формате доступно большое количество статей, связанных с рассматриваемой темой.
Маракушев С.А., Белоногова О.В. // Вестник Моск. Универ. 2010а. Сер. Химия. Т.51. С. 254-260.
Маракушев С.А., Белоногова О.В. Эволюция архаической хемоавтотрофной системы фиксации двуокиси углерода в гидротермальных системах. // Доклады Академии Наук. 2010б. №2. С.262-268.
Bada J.L. 2004. How life began on earth: a status report. // Earth Planet Sci Lett. V.226. P.1–15.
Baltscheffsky H. Inorganic pyrophosphate and the origin and evolution of biological energy transformation. In: Buvet R., Ponnamperuma C., editors. Chemical evolution and the origin of life. North-Holland Biomedical Press; Amsterdam, The Netherlands: 1971.
Cody G.D, Boctor N.Z, Filley T.R, Hazen R.M, Scott J.H, Sharma A, H S, Yoder H.S., Jr Primordial carbonylated iron–sulfur compounds and the synthesis of pyruvate. // Science. 2000. V.289. P.1337–1340.
Cody G.D., Boctor N.Z., Hazen R.M., Brandes J.A., Morowitz H.J., Yoder H.S. Geochemical roots of autotrophic carbon fixation: hydrothermal experiments in the system citric acid, H2O–(7FeS)–(7NiS). // Geochim. Cosmochim. Acta 2001. V.65. P.3557–3576.
Cody G.D., Boctor N.Z., Brandes J.A., Filley T.R., Fallick A.E., Ilich M., Russell M.J. . // Econ. Geol. 1991. V.86. P.847–861.
Fuchs G., Stupperich E. Evolution of autotrophic CO2 fixation. In Schleifer KH, Stackebrandt E. ed; Evolution of Prokaryotes, FEMS Symposium. 1985. No. 29. London: Academic Press. P.235–251.
Fontecave M. Iron-sulfur clusters: ever-expanding roles // Nature Chemical Biology. 2006. V.2. P.171-174.
Hazen R.M., Yoder H.S. Assaying the catalytic potential of transition metal sulfides for abiotic carbon fixation. // Ceochim. Cosmochim. Acta. 2004. V.68 P.2185–2196.
Corliss J.B. Hot springs and the origin of life. // Nature. 1990. V.347. P.624.
Di Gulio M. The universal ancestor lived in a thermophilic or hyperthermophilic environment. J. Theor. Biol. 2000. V.203. P.203–213
Dörr M. et al. Eine mögliche präbiotische Bildung von Ammoniak aus molekularem Sickstoff auf Eisensulfidoberflächen. // Angew. Chem. 2003a. V.115. P.1579–1581.
Dörr M., Kassbohrer J., Grunert R., et al. A possible prebiotic formation of ammonia from dinitrogen on iron sulfide surfaces. // Angew. Chem. Int. Ed. 2003b. .42. P.1540–3.
Kelley D.S. et al. . Nature. 2001. V.412. P.145–149;
Kelley D.S., Baross J.A., Delaney J.R. Volcanoes, fluids, and life at mid-ocean ridge spreading centers. // Annu Rev Earth Planet Sci 2002. V.30. P.385–491
Marakushev S.A, Belonogova O.V. The parageneses thermodynamic analysis of chemoautotrophic CO2 fixation archaic cycle components, their stability and self-organization in hydrothermal systems. // J. Theor. Biol. 2009. V.257. P.588-597.
Martin W., Russell M. 2003. On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. // Phil. Trans. R. Soc. Lond. B V.358. P.59–85.
McCollom T.M., Ritter G., Simoneit B.R.T. Lipid synthesis under hydrothermal conditions by Fischer-Tropsch-type reactions. // Origins Life Evol Biosphere 1999. P.29. P.153–166.
Hafenbradl D., Keller M., Wächtershäuser G., Stetter K.O. Primordial amino acids by reductive amination of α-oxo acids in conjunction with the oxidative formation of pyrite. // Tetrahedron Lett. 1995. V.36. P.5179–5182.
Heinen W., Lauwers A.M. Organic sulfur compounds resulting from the interaction of iron sulfide, hydrogen sulfide and carbon dioxide in an anaerobic aqueous environment. // Orig. Life Evol. Biosph. 1996. V.26. P.131–150
Hennet R.J.C., Holm N.G., Engel M.H. Abiotic synthesis of amino acids under hydrothermal conditions and the origin of life: a perpetual phenomenon? //
Naturwissenschaften 1992. V.79. P.361–3655.
Holm N.G., Anderson E.M., Hydrothermal systems. In: Brack, A. (Ed.), The Molecular Origin of Life; Assembling Pieces of the Puzzle. Cambridge University Press, Cambridge. 1998. P.86–99.
Holm N.G., Charlou J.L. Initial indications of abiotic formation of hydrocarbons in the Rainbow ultramafic hydrothermal system mid-Atlantic Ridge. Earth Planet Sci. 2001. V.191. P.1–8.
Huber C., Wächtershäuser G. Activated Acetic Acid by Carbon Fixation on (Fe,Ni)S Under Primordial Conditions. // Science. 1997. V.276. P.245-247.
Huber C., Wächtershäuser G. G. Peptides by activation of amino acids with CO on (Fe,Ni)S surfaces and implications for the origin of life. // Science. 1998. V.281. P.670–682.
Huber C., Wächtershäuser G. Primordial reductive amination revisited. // Tetrahedron Lett. 2003. V.44. P.1695–1697.
Lindahl P.A. Stepwise evolution of nonliving to living chemical system. // Origins Life Evol. Biosphere. 2004. V.34. P.371–389.
Leman L.J, Orgel L.E, Ghadiri M.R. Amino acid dependent formation of phosphate anhydrides in water mediated by carbonyl sulfide. // J. Am. Chem. Soc. 2006. V.128. P.20–21.
Leman L., Orgel L., Ghadiri M.R. Carbonyl sulfide-mediated prebiotic formation of peptides. // Science. 2004. V.306. P.283–286.
Ljungdahl L.G. A life with acetogens, thermophiles, and cellulolytic anaerobes. // Annu. Rev. Microbiol.2009. V.63. P.1–25.
Macleod, G., McKeown, C., Hall, A.J. and Russell, M.J. Hydrothermal and ocean ic pH conditions of possible relevance to the origin of life. // Origins of
Life and Evolution of the Biosphere. 1994. V.23. P.19-41.
McCollom T.M. Reduction of aqueous CO2 to organic compounds at hydrothermal conditions: Does it work? Presented at the First Astrobiology Science Conference, Ames Research Center, Mountain View, California, 2000. April 2-5.
Morowitz H.J., Kostelnik J.D., Yang J., Cody G.D. The origin of intermediary metabolism. Proc. Natl. Acad. Sci. USA 2000. V.97. P.7704–7708.
Nisbet E.G. Fowler. C.M.R. Some liked it hot. // Nature. 1996. V.382. P.404-405.
Nisbet E.G., Sleep N.H. The habitat and nature of early life.// Nature 2001. V.409 P.1083–1091.
Orgel L.E. // PLoS Biol. 2008. V.6: e18, 10.1371/journal.pbio.0060018.
Pereto J.G., Velasco A.M., Becerra, A., Lazcano, A. Comparative biochemistry of CO2 fixation and evolution of autotrophy. // Int. Microbiol. 1999. V.2. P.3–10.
Proskurowski G., Lilley M.D., Seewald J.S., Früh-Green G.L., Olson E.J., Lupton J.E., Sylva S.P., Kelley D.S. . Science. 2008. V.319. P.604–607.
Ragsdale S.W., Pierce E. Acetogenesis and the Wood-Ljungdahl pathway of CO2 fixation. // Biochim. Biophys. Acta. 2008. V.1784. P.1873–98.
Rao P.V. Holm. R.H. Synthetic analogues of the active sites of iron-sulfur proteins. // Chem. Rev. 2004. V.104. P.527-559
Russell M.J., Arndt N.T. Geodynamic and metabolic cycles in the Hadean. // Biogeosciences. 2005. V.2. P.97–111.
Russell M.J. First life // American Scientist. 2006. V. 94. P.32-39.
Russell M.J., Daniel R.M., Hall A. . // Terra Nova 1993. V.5. P.343–7.
Russell M.J., Daniel R.M., Hall A.J., et al. A hydrothermally precipitated catalytic iron sulphide membrane as a first step toward life. // J. Mol.
Evol. 1994. V.39. P.231–43.
Russell M.J., Hall A.J., Cairns-Smith A.G., Braterman P.S. // Nature. 1988.V.336. P.117.
Russell M.J., Hall A.J., Mellersh A.R. On the dissipation of thermal and chemical energies on the early Earth: The onsets of hydrothermal convection,
chemiosmosis, genetically regulated metabolism and oxygenic photosynthesis, in Ikan, R., ed., Natural and Laboratory-Simulated Thermal Geochemical Processes:
Dordrecht, Kluwer Academic Publishers, 2003. P.325-388
Russell M.J. The alkaline solution to the emergence of life: energy, entropy and early evolution // Acta Biotheoretica. 2007. V.55. No. 2. P.133-79.
Russell, M.J., Hall A.J. The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH
front. // J. Geol. Soc. (London) 1997. V.154. P.377–402.
Russell M.J., Hall A.J. From geochemistry to biochemistry: Chemiosmotic coupling and transition element clusters in the onset of life and photosynthesis // The Geochemical News. 2002. V.113. P.6-12.
Russell M.J., Martin W. The rocky roots of the acetil-CoA pathway. // Trends Biochem. Sci. 2004. V.29. P.358–363.
Russell M.J. The alkaline solution to the emergence of life: energy, entropy and early evolution. // Acta Biotheor. 2007. V.55. P.133–179.
Schidlowski M., Hayes J. M. Kaplan I. R. Isotopic inferences of ancient biochemistries: carbon, sulfur, hydrogen and nitrogen. In: Earth earliest biosphere (ed. Schopf J.W). 1983. P.149-186.
Seewald J. S., Zolotov Yu. M., McCollom T. Experimental investigation of single carbon compounds under hydrothermal conditions. // Geochimica et Cosmochimica Acta. 2006. V.70. P.446–460.
Smith E., Morowitz H.G. Universality in intermediary metabolism. // Proc.Natl. Acad. Sci. USA 2004. V.101. P.13168–13173.
Stetter K. O. Hyperthermophilic procaryotes. // FEMS Microbiology Reviews. 1996. V.18. P.149-158.
Taylor P, Rummery T.E, Owen D.G. Reactions of iron monosulfide solids with aqueous hydrogen sulfide up to 160C. // J. Inorg. Nucl. Chem. 1979. V.41. P.1683–1687.
Thauer RK. A fifth pathway of carbon fixation. // Science. 2007. P.318. P.1732–1733.
Von Damm K.L. Chemistry of hydrothermal vent fluids from 9°-10°N, East Pacific Rise: "Time zero", the immediate posteruptive period. // Journal of Geophysical Research 2000. V.105B. P.203-11,222.
Wächtershäuser G. An all-purine precursor of nucleic acids. // Proc. Natl Acad. Sci. USA. 1988a; V.85. P.1134–1135.
Wächtershäuser G. // Microbiol. Rev. 1988c. V.52. P.452–484.
Wächtershäuser G. Evolution of the first metabolic cycles. // Proc.Natl. Acad. Sci. USA 1990. V.87. P.200–204.
Wächtershäuser G. // Phil. Trans. R. Soc. B. 2006. V.361. P.1787-1808
Wächtershäuser G. Groundworks for an evolutionary biochemistry: the iron–sulfur world. // Prog. Biophys. Mol. Biol. 1992. V.58. P.185–201.
Wächtershäuser G. Pyrite formation, the first energy source for life: a hypothesis. // Syst. Appl. Microbiol. 1988b. V.10. P.207–210.
Wächtershäuser G. // Pure & Appl. Chem. 1993 V. 65, P. 1343-1348,
Wächtershäuser G. The origin of life and its methodological challenges. // J. Theor. Biol. 1997. V.187. P.483–494.
Yamagata Y., Watanabe H., Saitoh M., Namba T. Volcanic production of polyphosphates and its relevance to prebiotic evolution. // Nature. 1991. V.352. P.516–519.
de Zwart I..I, Meade S.J., Pratt A.J. 2004. Biomimetic phosphoryl transfer catalysed by iron(II)-mineral precipitates. // Geochim Cosmochim Acta V.68. P.4093–4098.
Статьи, связанные с темой.
Минеральные корни современных ферментов
Как мог выглядеть архаичный метаболизм?
Реликтовые составляющие современного метаболизма. С чего он мог начаться?
Список публикаций >>
Тест: А не зомбируют ли меня? Тест: Определение веса ненаучности
| Обнаружен организм с крупнейшим геномом Новокаледонский вид вилочного папоротника Tmesipteris oblanceolata, произрастающий в Новой Каледонии, имеет геном размером 160,45 гигапары, что более чем в 50 раз превышает размер генома человека. | Тематическая статья: Формирование субъективного опыта в онтогенезе |
Рецензия: Работы А.Иваницкого, комментарии | Топик ТК: Свойства осознанного внимания и циклы |