Алгоритмы сознания 2
Для входа в область понимания о “сознании” и “подсознании”, да и вообще “про мозг” есть дежурная статья с предельно наглядным показом, и внизу – рекомендованные ссылки на глубокое погружение. Последующий же текст во многом морально устарел, но в нем много полезного и интересного.
В попытках создать ИИ разработчиками моделей было потрачено неизмеримое количество времени, по большей части совершенно бесплодно потому, как взятое направление заходило в беспросветный тупик (например - "экспертные системы"). Эти потери времени были результатом, прежде всего, стремлений сделать разум, не повторяя природный (что подразумевает доскональное его изучение и чего нет до сих пор), а сразу "более эффективно", "другим путем", более "логичным". При этом казалось до степени убежденности, что интуитивно понятно, что нужно делать, чтобы задуманная конструкция при усложнении непременно начала проявлять качества разумной, но без понимания того, а в чем именно состоит эта "разумность".
На самом деле, тот единственный пример реализации разума, что дан природой в огромном множестве вариантов исполнения, но реализующий вполне четкие принципы механизмов адаптивности, использует все то необходимое и достаточное, что и представлено в природных адаптивных системах. И от этих принципов не отбросить ничего, кроме особенностей реализации, несмотря на, казалось бы, явную избыточность некоторых природных решений, хотя любая степень такой "избыточности", чаще всего, оказывается обусловленной определенной ее необходимостью. В потрясающей целесообразности природных механизмов легко можно убедиться, рассматривая уже хорошо изученные системы, например, систему слуха и вестибулярного аппарата. Стоит только разобраться, что и как для этого использовано и становится ясно, что ничего лишнего тут нет, а все организовано удивительно гармонично и единственно верно для использования в данных условиях.
Уверен, что, не разобравшись досконально с принципами природной реализацией адаптивного поведения и не имея никакого другого примера, никто не сможет придумать нечто лучшее, тем более, не понимая основной цели появления разума, а ориентируясь на его внешние проявления. И это - вполне обоснованное утверждение хотя бы тем, что человек оказывается не в состоянии придумать нечто принципиально новое, не имея доступных наблюдению аналогов (правда это утверждение, очевидно далеко не для всех :) ).
Поэтому, в данной статье, будет сделан упор на описание алгоритмов, использованных в природной реализации адаптивного поведения в их иерархии эволюционного последовательного появления. В этой иерархии все самые древние механизмы продолжают эффективно выполнять свои функции на уровне функциональности, характерной для данного этапа эволюции (см. Эволюция механизмов сознания).
Фактических данных по таким механизмам накоплено огромное количество, но подчас, без обобщения в виде системы.
Надо сказать, что в реальных нейронных сетях (а мозг представлен не одной сетью, а иерархией локальных сетей, созревающих строго поочередно в соответствии с последовательностью эволюционных усложнений) не используются точно подобранные весовые коэффициенты связей, хотя они в некоторых пределах и меняются. В мозге, для организации адаптивности звеньев поведенческих цепочек, используется установление все новых возбуждающих и тормозных связей (и отмиранее старых, давно не активировавшихся) при обучении, а на каждый распознаватель-нейрон не возлагается излишне сложная задача. (См. работы: Физиология центральной нервной системы и Проблема переработки информации в зрительной системе лягушки)
Это утверждение выглядит не однозначно, учитывая, что в рецепторной и эффекторной части нейронные отростки конкурируют между собой за соединение с мишенью и остаются те, что проявляют наибольшую импульсную активность, а синапсы тем более "укрепляются", чем чаще активность нейронов по обе их стороны (см. Формирование синапсов). Но при таком усилении проводимости синапса она стремится к ее нормальному для установленной связи максимальному значению, а не точной подстройке коэффициента синаптической передачи. В отличие от первичных рецепторных и эффекторных областей мозга (эволюционно древние структуры), более молодые зоны позволяют ассоциировать связи там, где это диктуется необходимостью новой адаптации, сколь угодно гибко изменяя функцию распознавания при изменении жизненного опыта хоть на противоположные по отношению к предшествующим поведенческие реакции - за счет использования признаков иерархии контекста распознавания.
В более молодых зонах, функции отдельных распознавателей могут выполнять целые участки сети (ансамбли возбуждения текущего восприятия в текущих условиях).
Сегодня существует много вариантов детально разработанных моделей "нейронных сетей", которыми пытаются имитировать реальные механизмы образования связей в мозге (например, см. Неокогнитрон Фукушимы), основанных на подборе весовых коэффициентов связей (что для природной реализации не характерно). Многие разработчики искренне считают, что такими сетями моделируются любые функции реальных механизмов мозга. Распространенное заблуждение, что если "правильно" изготовить и обучить сеть, то она сможет размышлять, давая ответы и как-то действуя, основываются на элементарном не понимании, что такое "размышлять" [249].
Нейронные сети мозга, сформированные из распознавателей, обладают далеко не только распознавательными функциями по принципу распознал-среагировал, как это организовано у насекомых (но и у насекомых распознавание - уже контекстно зависимо). Вот перечень наиболее важного из того, что призваны осуществлять механизмы мозга.
1. "Оперативная память" - то, о чем обычно говорится как об осознанном воспоминании (реверберирующие ансамбли возбуждения, только один из которых в данный момент времени осознается) в одном из трех режимов осознания: 1) отслеживание наиболее актуальных событий [249], 2) переключение на более желательное в данной специфике условий действие из набора уверенных продолжений, 3) прерывание текущей цепи действий для творческого нахождения более желаемого варианта в данных условиях.
В каждый данный момент времени могут неосознаваемо выполняться множество различных поведенческих цепочек за счет ранее отработанного с помощью осознания опыта такого поведения (цепочек, в которых каждая последующая фаза запускается концом отработки предыдущей, и которые могут состоять из других цепочек поведения - в качестве отдельных фаз). Внимание сопровождает ту фазу из них, в которой возник новый, требующий переадаптации элемент (обычно - признак новых условий), важный для результата отработки данной цепочки. Т.е. внимание переключается постоянно на ту фазу всех активных цепочек, в которой произведение новизны на значимость оказывается наибольшее.
При этом актуальная фаза цепочки поведения замыкается через ассоциированную значимость переключателем, связывающем актуальную активность так, что активность становится самоподдерживающейся и возникший образ данного звена восприятия-действия субъективирован связанной с ним значимостью, делая его в таком качестве доступным продолжительному вниманию для механизма сознания (фокус сознания, исследованный Иваницким А.М.). Чем более актуален образ, чем более актуальна не решенная проблема новой ситуации, тем более стабильно поддерживается такая реверберация образа, вплоть до характера доминанты [9].
Для обеспечения нормального функционирования этой системы сознания, для гашения накопившихся активностей, остающихся после касания сознанием, служит не только взаимное латеральное торможение, но и "функциональная система" сна, но лучше назвать ее контекстом поведения перехода ко сну (потому, что как и другие эмоциональные контексты она в основе регуляции используется специфические нейромедиаторы).
2. Переключения эмоциональных контекстов поведения [106], обеспечивающее оптимальный стиль поведения в данных условиях и определяющий преимущественный характер установления адаптирующих связей: блокирующий нежелательное направление цепочки поведения (- отрицательная значимость эмоционального контекста) или стимулирующий ее (- положительная значимость эмоционального контекста).
3. В качестве цепочек поведения организована и мыслительная деятельность - последовательность переключения внимания по ранее акуальным фазам цепочек, где эффекторами выступают цепи возбуждения и торможения, обеспечивающие переключение каналов восприятия - внимание.
Регулировка внимания используется для выбора более желательного
варианта поведения из уже уверенно известных по опыту вариантов поведения в данных
условиях или - для перехода к поисковому поведению нового варианта. Она следует осознанию наиболее актуального, имеющего
функцию отслеживать то, что в данный момент может потребовать адаптации, делая оценку по
шкале хорошо-плохо, - оценку значимости совершающегося в данных условиях (в
других условиях то же самое может иметь совершенно иную значимость).
Чтобы внести ясность в понимание того, что означает слово "сознание", привожу ссылку на определение: Определение сознания.
4. Целеобразующий прогноз при осознании - на основе "опережающих возбуждений" [33], позволяющий предсказать наиболее вероятный итог действия и тем самым выбрать наиболее подходящее действие.
5. Эволюционно создалась новая кора - как место локализации специализирвоанных распознавателей примитивов восприятия (в отличие от единой сети, мозг построен как иерархия многих сетей, начиная с самых элементарных). Каждый новый слой этой коры образовывался в виде все более усложняющихся примитивов и обеспечивает разделени распознавателей для различных стилей поведения (эмоций) с помощью специфичности синапсов к определенному нейромедиатору [22].
По сравнению с распознаванием на уровне сети третичных зон [223] мозга (ансамблей возбуждения звеньев поведенческих цепей) распознавание примитивами восприятия обеспечивает неизмеримо более надежный и быстрый результат при большей простоте организации (не требуется дообучения огромного числа ранее установленных связей, необходимое при каждом новом обучении). На ранних этапах развития организма заготавливаются все возможные примитивы, с которыми сталкивается восприятие в этот период, после чего это и определяет эффективности интеллектуальных способностей. Лишение восприятия стимулов в соответствующие критические периоды развития лишает психику возможности использовать признаки этих стимулов.
Как уже говорилось, все это организуется в виде эволюционно-последовательно созревающих, функционально усложняющихся слоев распознавателей (см. Организующий принцип функции мозга). Думаю, что моделирование адаптивного поведения должно использовать структурные элементы и принципы, используемые в нервной системе любого существа, даже организованного не сложнее, чем насекомое. В последовательности эволюционного усложнения и созревания и будем их конкретизировать.
Первые сочетания в распознавателях примитивов восприятия не используют адаптивность установления возбуждающих или тормозных связей (обучение с использованием механизмов сознания: системы оценки результата распознавания, "с учителем").
Порог срабатывания определяется фоном подтормаживающего окружения (взаимное торможение всех элементов сети) и медиаторной спецификой синапсов. Порог, как правило, значительно больше единицы действия проводящего синапса, и распознаватель может не сработать от одного рецептора. Но когда возбуждение через порог прорвется, то распознаватель сам начинает подтормаживать соседей.
В модели, имитирующей уровень развития распознавателей "до учителя", необходимо реализовать механизм образования последовательно созревающих слоев распознавателей примитивов восприятия со все более усложняющейся функцией распознавания. Не важно, как именно это будет реализовано и насколько похоже на природную реализацию, важно чтобы были получены наборы примитивов, коллекции распознавателей тех образов восприятия, которые предъявлялись в период развития слоев распознавателей. В пояснении описаны реализуемые принципы.
Стоит отметить, что для разных рецепторных полей этот
алгоритм в природе реализуется несколько по-разному, а не универсально, что
обеспечивается определенным предопределением развития. Особенно много нюансов
при формировании распознавателей зрительного восприятия.
В общем же плане, каждый последующий слой - эволюционно обусловлен и должен следовать общему принципу экспрессии генов и т.п. механизмам неследуемости, хотя, возможно, что специального гена на каждый слой и не существует. Чисто же функционально, из всей совокупности известных данных вырисовывается следующая картина.
Когда есть предшествующие рецепторы, и они проявляют активность во время восприятия, то к источникам ближайшей активности от последующего слоя нейронов начинают ветвится дендриты (это - общий принцип). После чего появляется возможность установления связи по стандартному условию ее возникновения [20]. Когда таким образом сформируется слой примитивов, то он уже начнет проявлять активность в качестве рецептора для последующего. И т.п.
Эта картина дополняется тем, что пока у созревшего нейрона нет связей, он проявляет пейсмеккерную активность [7], что и создает условия для фиксации связей с теми из предшествующих рецепторных нейронов, который в то же время проявили активность. За это время профиль активности рецепторов может несколько меняться (насколько это критично и как образуется наиболее общее - предлагается посмотреть в моделе). Допускаю, что есть некоторые механизмы, например, 1. поддерживающих возникшую активность связки предшественников и нового нейрона даже после исчезновения стимула, 2. тормозящие соседние мешающие образы, 3. следовые последействия ухудшающие условия образования новых сочетаний и т.п. [284]. Главное, что для целей алгоритмической прикидки в конкретной реализации можно осуществить наиболее удобные поддерживающие механизмы.
Несмотря на то, что нейрон имеет до 10000 потенциальных входов, распознаватели не "играют" на тонких изменениях уровней потенциалов порога срабатывания (порог у мембраны довольно груб да еще зависит от многих факторов), у них другой, более надежный принцип распознавания момента срабатывания (принципиальное отличие от модели персептрона). Важно то, что каждый вход имеет определенный информационный смысл, заключающийся в том, что он идет от определенного предшествующего рецепторного выхода (каким может быть предшествующий распознаватель или непосредственно рецептор). Сочетание таких активных смыслов, часть из которых возбуждающая, а часть - тормозная, составляет профиль распознавания.
При этом нейрон - контекстный распознаватель, т.е. смысл его активности зависит от его расположения среди других колонок в определенной зоне. Поэтому профиль распознавания получается достаточно однозначен (хотя и не без ошибок, "иллюзий") хотя если бы не было контекста - локализации, то данное количество сочетаний возбуждающих и тормозных активностей на входе было бы характерно для многих разных профилей. Например, если представить 7-сегментую матрицу для показа чисел и букв в качестве аналогии, то профиль числа 0 точно такой же как профиль буквы О, но т.к. распознавание чисел локализуется в другой зоне, чем букв, то профиль оказывается однозначным.
Вне зависимости от способа реализации, важно, что принцип использования усложняющейся иерархии примитивов - общий для любых природных реализаций. Главное, что достигается: формирование наиболее актуальных для данной особи и ее окружения наборов примитивов (из бесконечного количества вообще возможных), которые будут использованы как рецепция корректируемых сознанием распознавателей.
Для формирования распознавателей "с учителем" (т.е. при участии сознания и появлении коммутатора фокуса сознания (гиппокамп) - внимания с использованием системы значимости [125]), опытом сопоставления желаемого и получаемого нарабатываются критерии ошибки распознавания, - не соответствие желаемому результату, где желаемость - оценка системой позитивного результата своего акта действия - как достигающего цели выполнения этого акта. Сигнал ошибки провоцирует установление тормозных связей с пусковым распознавателем акта действия. Предоставляется соответствующая модель (2) эффекторного распознавателя (всякий распознаватель является в принципе эффекторным по отношению к предыдущим, что и обуславливает выполнение мыслительных цепочек действия так же как и мышечных) для визуального экспериментирования, в пояснении которой приводится ее исходный код для свободного использования.
Обе представленные модели идеализированы. В них не
учитываются предопределенности, определяющие специфику формирования
распознавателей в слоях, а так же то, что связи образуются не моментально и не
сразу полновесно, что влияет на результат обучения. В модели, где используется
результат оценки для научения, этот результат получается не в самой модели, а
используется оценка экспериментатора. Но ниже будет показан принцип такой
оценки самой системой, и этот принцип проиллюстрирован схематично.
То, что подобные распознаватели могут ошибаться во множестве случаев когда составляющие образ признаки будут оказываться в несколько ином порядке, но распознаватель этого не будет замечать пока реальность не даст по башке (несоотвествием желаемого и получаемого), требуя дополнительного обучения. С другой стороны это позволяет очень быстро делать выводы даже в случаях, несколько отличающихся от запомненной модели.
Так, по рзелульаттам илссеовадний одонго анлигйсокго унвиертисета, не иеемет занчненя в кокам пряокде рсапожолены бкувы в солве. Галвоне, чотбы перавя и пслоендяя бквуы блыи на мсете Осатьлыне бкувы мгоут селдовтаь в плоонм бсепордяке, все-рвано ткест чтаитсея без побрелм. Пирнчиной эгото ялвятеся то, что мы не чиатемкдаужю бкуву по отдльенотси, а все солво цликеом. Т.е. распознаватель слова имеет настройку на первую и последнюю буквы, а так же на наличие других букв, но уже вне какого-то порядка.
И даже вот такое распознается при беглом просмотре:
D94НН03 С006Щ3НN3 П0К4ЗЫ8437, К4КN3 У9N8N73ЛЬНЫ3 83ЩN М0Ж37 93Л47Ь Н4Ш Р4ЗУМ! 8П3Ч47ЛЯЮЩN3 83ЩN! СН4Ч4Л4 Э70 6ЫЛ0 7РУ9Н0, Н0 С3ЙЧ4С Н4 Э70Й С7Р0К3 84Ш Р4ЗУМ ЧN7437 Э70 4870М47NЧ3СКN, Н3 З49УМЫ84ЯСЬ 06 Э70М. Г0Р9NСЬ. ЛNШЬ 0ПР393Л3ННЫ3 ЛЮ9N М0ГУ7 ПР0ЧN747Ь Э70
Подобное нужно читатаь как можно меньше напрягая внимание и сознание. Тогда раскручивается наиболее накатанная цепь понимания. При обострении же внимания к фрагментам выясняется их новизна и не соотвествие ранее наработанным шаблонам. Для нахождения важных отличий и нужно сознание.
О том, каковы могут быть реальные затраты времени на формирование распознавателей, можно судить по статье В мире символов:
...детям требуется несколько лет, чтобы полностью постичь природу изображений. Джон Флейвелл (John H. Flavell) из Стэнфордского университета обнаружил, что до четырех лет многие дети полагают, что если перевернуть нарисованный стакан с попкорном, то его содержимое высыплется. Они также убеждены, что фотоснимок должен меняться по мере того, как в реальном мире происходят изменения с тем, что на нем запечатлено. Подобные заблуждения свидетельствуют о том, что малыши еще не вполне осознают некоторые аспекты взаимосвязи между предметом и его изображением.
В реальных алгоритмах возможно не использовать этапы формирования примитивов восприятия. Достаточно было бы, изучив их вариации и назначение, сразу для каждого вида восприятия заготовить такие примитивы - как готовые функциональные элементы. Но их количество само по себе огромно и всеобъемлюще перекрывает все прогнозируемые возможные области восприятия. Так, если нужно, чтобы система очень быстро могла "на вскидку" прикидывать количества предметов в поле зрения, то нужно заготовить все распознаватели для всех количеств предметов заранее. Или, если системе предстоит уметь быстро вычислять "в уме", то она должна быть снабжена примитивами всех таких вычислений (что делается и в современных процессорах с помощью микропрограмм вычислений).
Можно также заготовить и некоторые поля распознавателей более высокого уровня интеграции, использующие различные типы рецепторов. Но их общее число окажется еще больше...
Без использования такого предварительного распознавания (коллекции примитивов), с попыткой нагрузить этой функцией "основную сеть", невозможно будет получить сколько-то приемлемую эффективность и надежность адаптивной системы даже для насекомоподобной модели с функциональностью по типу: распознал-ответил.
Но, кроме этого, существуют еще и распознаватели "третичных зон" (где сходятся все рецепторные распознаватели и имеется доступ к эффекторным командным распознавателям), призванные интегрировать выходы более простых распознавателей с выходом на бессознательное (автоматическое) реагирование.
Если этим заканчивается сложность организации нервной системы у насекомых, то для эффективной адаптации к новым условиям это является только самым началом организации :) Т.е. в этом начале мы имеет только коллекции (поля) обученных распознавателей различной сложности, часть из которых непосредственно организуют ответные действия.
Далее предстоит сформулировать алгоритмические модели, позволяющие настраивать распознаватели, имеющие выходы на поведенческое действие так, чтобы они приводили к разному поведению при учете специфики разных условиях, т.е. поведение - в зависимости от специфики условий.
Так как условия могут меняться во время выполнения действий, то любое действие дробится на элементарные фазы, начало каждой из которых запускается успешным концом предыдущей и/или пусковым стимулом [308]. Так организуются реальные цепочки поведенческих актов. Сами по себе такие цепочки могут выполняться автоматически (без контроля сознанием), но если во время выполнения возникает новизна в условиях, то эта новизна должна давать возможность оценить возможные результаты при таком новом повороте событий, выбрать наиболее желательный вариант и скорректировать фазы текущей реакции или запустить цепь уже другой реакции. Кроме того, чтобы не запускать наиболее привычную цепочку из многих существующих при критичном изменении условий, необходима работа механизма, оценивающего возможный результат действий - прогноз, что является функцией механизма сознания.
Это реализуется с помощью общего для всех реакций и для всего восприятия механизма переключения "внимания" и оценки значимости, обеспечивающего фокусировку (подключение) механизма осознания на экстремально актуальном. Надо сказать, что условия практически никогда не бывают идентичными и можно говорить только о степени их новизны, как и степени значимости возможных, прогнозируемых прежним опытом, оценок значимости. Поэтому внимание обслуживает ту поведенческую цепочку, которая соответствует наибольшему произведению новизны на значимость (см. Механизм ориентировочного рефлекса ) и на это время теряется контроль сознания над всеми другими процессами, которые продолжают свое выполнение автономно (в том числе мыслительные цепочки, продолжающиеся бессознательно. Мыслительные процессы - те же поведенческие цепочки, только вместо управления мышцами управляется переключениями существующих активных ансамблей возбуждения мозга и активация ассоциативно связанных с ними).
Каждая из фаз поведенческой цепочки может быть приостановлена воздействием внимания и зациклена связью с ее выхода на вход через ассоциированные распознаватели системы значимости, порождая субъективный образ "оперативной памяти" этой фазы. Если это фаза мышечного действия, то в этом положение движение останавливается и поддерживается реверберацией, если это фаза "мысленной" цепочки памяти, то образ "стоит" в сознании. Кроме цепочек, выполняющихся автономно в фоне осознания, такие остановленные мгновения (в том числе впечатления от текущего восприятия, оставленные сознанием из-за более важного) могут продолжать реверберировать пока не будут погашены взаимным торможением или во время сна.
Модель сознания предполагает создание системы
распознавателей, способной выбирать наиболее значимый вариант поведения в
сложившихся условиях и в зависимости от результата его реализации фиксировать
связи всех сопутствующих образов восприятия с программами действия так, что нежелательный
результат будет образовывать тормозные связи, а желательный - возбуждающие связи
с пусковыми элементами поведения. Таким образом, в последующем, в данной
ситуации уже даже без контроля сознания реакция пойдет в желательном
направлении автономно. Если прошлый опыт не предоставляет успешных прогнозов,
то реализуется, так же постоянно оптимизирующаяся жизненным опытом, общая
программа поиска благоприятного реагирования.
Понятно, что в таком случае сознание - не нечто локализованное в какой-то части мозга, а его составляют все участвующие в адаптивности составляющие. Соответственно, утрата какой-то из составляющих определенным образом означает утрату этой функциональности сознания.
Организация корректируемой осознаваемым опытом системы распознавателей с творческим продуцирование новых вариантов поведения - эволюционно наиболее
молодое образование в природной реализации, то, что мы обычно называем интеллектом или жизненным опытом.
Этим формируется система распознавателей в цепочках навыков творчества (поисковое поведение). Если
сформированный мозг оказывается лишенным наработанных цепей творческой адаптивности (лоботомия), то весь
закрепленный опыт поведения сохраняется (в том числе и опыт речевой коммуникации), но уже
не может быть модифицирован в зависимости от специфики
условий, что проявляется как отсутствие способностей приспособления к новому. Или же достаточно просто повреждения переключателя фокуса сознания (гиппокампа), чтобы система осознанной адаптивности не могла нарабатывать новый опыт и вообще называться осознаваемой (ведь повреждается механизм субъективности при зацикливании образов восприятия-действия).
Новейшие зоны могут отключаться во многих состояниях, например, при наркотических отравлениях, гипоксии, усталости. Часто они оказываются малоактивными в хорошо знакомых ситуациях, не предвещающих никаких новых переживаний. При этом не происходит запоминания цепочек происходящих событий. Состояние же острого и ясного внимания всегда сопровождается и четкими следами памяти о происходящем. Это и есть адаптивность психики в действии.
При параллельном выполнении многих поведенческих автоматизмов, которые обслуживают все, от восприятия до реализации всех элементов поведения в текущий момент, сознание подключается к той фазе того автоматизма, что имеет наибольшее произведение новизны-значимости, т.е. является наиболее актуальным для возможного вмешательства и корректировки.
С точки зрения алгоритма механизма сознания, возможны три типа развертывания событий.
1. Если моментальный прогноз от этой фазы не даст негативной оценки, превышающей допустимый в данных условиях порог, или этот прогноз не даст позитивной оценки для другого известного варианта поведения, превышающий допустимый порог риска, то сознание просто остается в режиме отслеживания текущих событий, переключаясь по цепи наиболее актуальных фаз и оставляя ленту памяти - воспоминания, которые могут вспоминаться.
По закону фиксации переживаемого, образуется цепочка псевдоавтоматизма последовательности воспоминаний, т.е. не участвующего в непосредственном поведении автоматизма, но формируемая точно так же как цепочка поведенческого автоматизма, т.е. укрепляясь от опыта к опыту сходного поведения.
Эта цепочка, в случае если сознание переводит свой фокус на нее - есть те самые "мысли", которые остаются доступными при воспоминании о событиях прошлого. Эта цепочка - есть "мысли", кинофильм впечатлений [249], что остаются доступными при воспоминании о событиях прошлого. Это - как бы взгляд на происходящее с точки зрения сознания настоящего, взгляд на то, что было определено сознанием прошлого. Понятно, что при каждом новом осознании прежняя цепочка модифицируется в соответствии с жизненным опытом нового сознания. Стоит что-то вспомнить из прошлого и воспоминания об этом становятся уже другими.
Можно не прибегать к специально выделенному понятию "псевдоавтоматизма", а просто сказать, что в ходе реализации параллельных автоматизмов с их фазами ассоциируются и сопутствующие образы творческого осознания в виде всплесков прогностических прикидок. Такие ассоциации развертываются при вспоминании о прошлых событиях как след сознания, отслеживавшего текущее восприятие-действие, след, доступный в виде цепочки впечатлений с ассоциированными ветвленими прогностических оценок - прикидок возможных последствий других вариантов с оценкой их допустимого риска. Чем больше задумывалось об этом в данный момент, тем более глубок этот след, иначе он просто не болеее, чем цепочка текущих впечатлений (оценок значимости) происходящего.
2. Если при отслеживании сознанием в несколько новых условиях (а они есть, раз сознание оказалось на данной фазе) прогноз выполнения действий не желателен, но есть уверенный опыт альтернативных действий, дающих более желаемый результат, то происходит переключение на него и фиксация, что в при такой новизне условий автоматизм имеет такое ветвеление.
3. Если в ходе прогноза появляется запрет на продолжение действий из-за ясного прогноза не желательного, а актуальность действия не настолько высока, чтобы ветвиться на слишком не уверенный вариант действия, а позволяет "подумать", то действие приостанавливается для творческой выработки продолжения с наиболее желательными последствиями. Точно так же, если даже при позитивном прогнозе возникает вариант с гораздо более желательным исходим, то так же действие может быть приостановлено для творческой корректировки [143].
Возникает противоречие (конфликт оценок) с тем, что ранее считалось наиболее приемлемым и было зафиксировано в виде автоматизма и тем, что возникло в ходе творческого поиска более желательного. Итог этого конфликта должен решить: или продолжить прежний автоматизм или начать выполнять с некоторым риском новый вариант поведения. При достаточно сильном расхождении проблемы не возникает, а вот в случае примерно равных оценок борьба конфликтов тем больше, чем больше важность ситуации, и это конфликт может быть довольно мучительным. Это то, что называют проявлением "воли", т.е. оценки осознанной в противовес с прежде закрепленной за данной фазой автоматизма оценкой прежних актов сознания. Т.е., фактически, это - борьба сам с собою из прошлого (более подробно о воле см. в статье Мотивация, Воля).
Сознание имеет дело со всеми доступными распознавателями восприятия, в том числе и распознавателями значимости - как текущего состояния организма по шкале хорошо-плохо. Первичными рецепторами системы распознавателей значимости (или просто системы значимости) являются рецепторы гомеостаза, болевые, термо и другие рецепторы аварийных ситуаций или избавления от них.
Частный случай поведенческого акта - формирование отдельных символьных распознавателей. Как правило, такие распознаватели представлены первоначально целым ансамблем возбуждения, непосредственно отражающим явление, а затем возможно формирование отдельного распознавателя, реагирующего на весь этот первоначальный ансамбль возбуждения и в последующем могущий заменять его. Уточнение связей с тем или иным ансамблем восприятия и является результатом оценки сознанием правильности распознавания или, точнее, успешности распознавания.
Сознание модифицирует активный профиль восприятия, связывая его с профилем распознавателей значимости (с учетом только действующего в данный момент эмоционального контекста и соответствующих ему синапсов, специфичных к определенному нейромедиатору). Для каждого из отдельных распознавателей профиля устанавливаются положительные связи в случае если есть уверенность в правильности распознавании ситуации (высокое значение произведения новизны на положительную значимость правильности распознавания), и отрицательные (высокое значение произведения новизны на отрицательную значимость правильности распознавания – т.е. детектор ошибки понимания ситуации – тоже нарабатывается личным опытом). При этом сочетание данного профиля с распознавателями положительной значимости стимулирует поведенческое действие, а отрицательной – препятствует ему. Побочным результатом последовательности таких актов, направленной на корректировку текущей цепочки поведенческой активности, является формирование сопутствующей цепочки воспоминаний, которая может запускаться с блокировкой выполнения связанными с ними фазами действий.
Понятно, что зоны распознавателей для обеспечения функции сознания
так же формируются от самых простейших примитивов к наиболее сложным.
Со стороны лобных долей в направлении переключателя осознанного внимания (гиппокампа) так же есть первичные, вторичные и третичные зоны. Наиболее древняя - обеспечивает автоматическую регуляцию «заученных» двигательных актов, поддержание общего мышечного тонуса, готовность периферического двигательного аппарата к совершению движений, перераспределение мышечного тонуса при движениях. Кроме того, она участвует в поддержании нормальной позы....В первичноймоторной коре (поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей....Вторичная двигательная кора (поле 6) имеет главенствующее функциональное значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. [334].
При этом лобные доли специализируются именно на участии в обработке новых ситуаций: Когда задача (назвать глагол, подходящий к визуально предъявленному существительному) была дана впервые, кровоток в лобных долях достиг высшего уровня. По мере знакомства испытуемых с задачей участие лобных долей почти сходило на нет. Когда предъявлялась новая задача, имеющая в целом сходство с первой, но не тождественная ей, кровоток в лобных долях несколько усиливался, но не вполне достигал своего начального уровня. Такое впечатление, что имеется строгое соотношение между новизной задачи и уровнем кровотока в лобных долях: он является самым высоким, когда задача является новой, самым низким, когда задача знакома, и промежуточным, когда задача является частично новой. [91].
Таким образом, рецептивным полем для формирования первичных зон лобных долей являются выходы распознавателей всех уровней и всех зон, но уже вторичные и третичные зоны модулируются контекстом текущих условий, как и в других каналах восприятия. Для длбных долей несомненна связь такого контекста опосредовано структурами гиппокампа, переключающего регулируемое звено цепи повеления. Управляется же переключение как сигналом новизны-значимости от ассоциативных зон мозга ("непроизвольное внимание"), так и со стороны самих лобных зон ("произвольное внимание").
В соответствии с принципом
двунаправленности связей, аксоны распознавателей зоны функции сознания проникают
во все слои рецепторных и эффекторных распознавателей (-командных нейронов),
обеспечивая их выделение вниманием и позволяя формироваться связям в этой
выделенной области между возбужденными элементами для специфичных к текущему
эмоциональному контексту медиаторов. Что и является сигналом оценки, пропорциональным
произведению новизны на значимость (положительную или отрицательную) для
текущего активного образа восприятия-действия, т.е. выделение медиатора,
характерного для данного эмоционального контекста, влияет по эффекту
"спиловера" (в переводе с английского означает перелив и
растекание) на синапсы распознавателей оценки новизна-значимость по
функции модуляции или перемножения (см. Нейрональная пластичность и Диффузная внесинаптическая нейропередача).
В отличие от неспецифической модуляции спиловера, есть нейроны-модуляторы, способные осуществлять операцию перемножения на уровне конкретного синапса: [155] .
Так как визуальную модель подобного взаимодействия разработать не просто (мягко сказано), то придется ограничиться мысленной моделью и схемной визуализацией.
Предположим, что в силу очень высокой облегченности образования "наследственной" связи между некоторым сочетанием рецепторных распознавателей и эффекторным распознавателем (-командным нейроном) установлена связь, образующая цепочку "рефлекторного" поведения. В раннем онтогенезе этот рефлекс срабатывал автономно по принципу распознал-среагировал.
При созревании распознавателя зоны механизмов сознания, он фиксирует профиль на своем входе в текущем эмоциональном контексте, где кроме признака условий данного движения присутствует и текущая оценка его непосредственных последствий - как сигнал распознавателей системы значимости, модулированный (функция перемножения) распознавателями новизны данного профиля возбуждения. Спонтанная активность созревшего распознавателя сочетается с таким профилем так, что в последующем при значении результата модуляции сигнала значимости новизной, он будет впредь активизироваться.
При этом, в случае отрицательной значимости, создаются условия для формирования тормозных связей в области сочетаний активного профиля восприятия с эффекторной фазой действия, а в случае положительной значимости - для формирования возбуждающих связей.
Теперь выход сформированного распознавателя будет способен активировать весь текущий профиль возбуждения даже при отсутствии реальных стимулов или же тормозить его даже если стимулы есть. После схожего восприятия весь образ будет возбужден и это возбуждение станет поддерживаться за счет влияния распознавателя зоны сознания. Кроме того, под его действием закрепляются связи от выходов распознавателей активного профиля на его вход так, что реверберация продолжается и после прекращения возбуждения распознавателя зоны сознания.
Этим регулируется возможность сознательного управления выделения восприятия вниманием: активируется тот распознаватель зоны сознания, где произведение новизны на значимость окажется наибольшим, при этом гася другие распознаватели (как это происходит во всех слоях распознавателей для контрастирования какого-то одного, наиболее актуального).
В последствии, при возникновении достаточно сходных условий для стимула, рефлекс начнет выполняться автономно, если не было установлено его блокировки сознанием ранее. Т.е. ранее безусловная реакция стала в данных условиях запрещенной. На уровне осознания, возбуждение мысленно еще до выполнения действия уже даст записанную в общем профиле оценку его результата, что может быть использовано как прогноз.
Точно так же сознательно (при наличии новизны в профиле условий) может корректироваться по результатам каждая из фаз поведенческой цепочки, что в последствии, позволяет получать прогностическую оценку до ее выполнения, просто вызывая мысленно сопутствующий образ. С другой стороны, появление уже оцененных системой значимости образов восприятия приводит к эффекту "понимания" - предвидению того, что они могут означать, к чему приводить в данных условиях на основе прошлого опыта.
Следует отметить, что данное представление - слишком
гипотетично. Так, одно из множества того, что требует уточнения: модуляторная
функция при перемножении сигнала новизны на значимость, возможно, реализуется с
помощью эффекта "cпиловер",
который достоверно используется в реальной нервной системе, обеспечивая
модуляцию синоптической проводимости диффузным распространением медиаторов.
Однако, это не мешает пониманию принципов функционирования
сознания, и модель способна помочь контрастировать направления исследования и
моделирования. В этом плане предполагается, что последующий текст сначала описательно, затем на примерах и схемах поможет создать это понимание :)
Итак, распознаватели зоны сознания не являются просто детекторами текущих профилей возбуждения и их символами. Они безотносительны в этом смысле к любым профилям: любой профиль текущего восприятия может оказаться в "фокусе внимания" этих распознавателей. Задача этих распознавателей - переключаться на тот профиль возбуждения из существующих, который имеет в данный момент наибольшее значение произведения новизны на значимость (значимость, связанную с прошлым опытом) в условиях соответствующей этому профилю выполняемой фазы поведенческой цепочки и, перебирая ассоциированные (похожие) цепочки схожего прежнего опыта, выбрать наиболее желательный (связанный с положительной значимостью) вариант продолжения поведения. Или, если нет таких аналогов, запустить общую поисковую стратегию (которая так же усложняется с опытом).
Т.е. для некоей ранее автономной поведенческой цепочки, критичная (новизна-значимость) фаза приостанавливается возбуждением распознавателей прежнего опыта и выбирается связанный с наиболее желанным исходом, который может запустить уже совсем другую цепочку поведения или продолжить прежнюю, после чего оценка результата породит новый жизненный опыт. Или если еще не было никакого предшествовавшего опыта, то запустит цепочку общей тактики на этот случай, опять-таки, после чего новый оцененный опыт уже появится.
Вот еще более схематичная интерпретация (за счет упрощения модели, конечно). Максимальное значение произведения новизны на значимость от всех текущих активных профилей восприятия-действия, возбуждает специфичный к этому профилю распознаватель зоны сознания и гасит остальные, выделяя доминирующий профиль для возможной коррекции текущей фазы поведения. Если существуют распознаватели, в которых элемент новизны вызывает возбуждение негативного последствия такого поведения, то возникает переключение на цепочку с более благоприятным исходом или включение общей стратегии выбора более желательного исхода. Если нет, то поведение продолжается. Результат этого продолжения в виде оценки системой значимости окажется связанным с данным распознавателем зоны сознания в виде нового опыта и одновременно связанным с самим профилем восприятия-действия или блокируя его для последующего или стимулируя.
Чисто умозрительный пример: если у улитки есть природный рефлекс ползти к издающему пищевой запах плоду, то она это делает всегда когда появляется запах, учитывая только степень своей сытости и вообще возможность ползти. Если на дороге возникнет новое для нее опасное препятствие (непосредственно не затрагивающее ее репцию аварийных состояний), она погибнет, несмотря на то, что в других условиях была только что научена избегать это препятствие, но там не было пищи (при наличии пищи предпочтение всегда отдается реакции избегания). Появление препятствия, которое в других условиях у нее связалось с опасностью (временное, <40 дней "обучение"), на пути к пище - очень значимая новизна, которая должна бы остановить существо, обладающее способностью осознавать критически важные фазы своего поведения.
В отличие от улитки, у высших животных связывание признаков опасности происходит не просто с программами избегания аварийных ситуаций, а с распознавателями нежелательных последствий (системы значимости) и при появлении этих признаков в других условиях, которые не были связаны с опасностью, вызывает появление компонента новизны (признаки необходимости внимания) и высокую отрицательную значимость, что должно обратить внимание на это при выполнении поведенческой цепочки и воспрепятствовать ее продолжению.
Основа алгоритма адаптации к новому в восприятии, если еще нет никакого опыта оценки последствий такова:
1. Новое вообще не замечается (не осознается в виду того, что произведение на нулевую значимость не приведет к активации сознания).
2. Некие последствия для организма возможны, и в таком случае уже есть ненулевое произведение новизна-значимость, что фокусирует внимания, после чего, в случае нежелательного результата с профилем текущего восприятия, несущем новое, связывается результат запуска общей программы избегания. В случае же положительного результата, с профилем связывается положительная значимость и в последующем такое бывшее новое уже будет как-то оценено и способно учитываться при обращении внимания.
Основа алгоритма адаптации к новому в восприятии, если уже есть связанные со знакомыми профилями текущего восприятия цепочки ответных действий такова:
1. Новизны нет, и ранее оцененная значимость без участия сознания будет включать наиболее желательную цепочку (прогностическое возбуждение с ассоциированной позитивной оценкой результата). Вот как это можно было бы проиллюстрировать схемно:
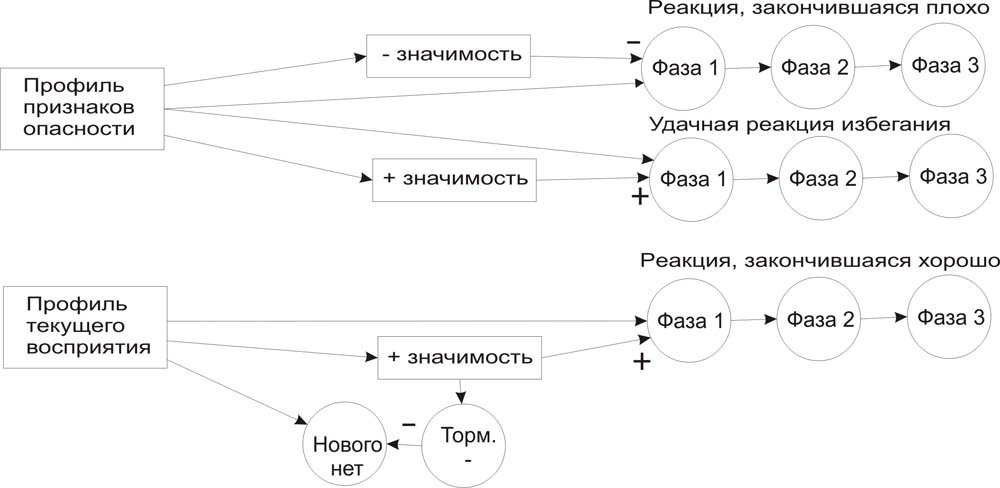
Основа алгоритма адаптации к новому в восприятии, если уже есть связанные со знакомыми профилями текущего восприятия цепочки ответных действий, но в ранее известном профиле появились признаки другого ранее известного профиля опасности такова:

Здесь срабатывает детектор нового потому, что в общем профиле восприятия появился дополнительный элемент, но еще нет отношения к такому сочетанию системы значимости. Однако, признаки опасности оцениваются старым распознавателем системы значимости. Высокая отрицательная значимость при новизне - сигнал того. что нужно приостановить текущую фазу реакции и заменить ее на ту известную, которая уже сформирована прежним опытом для такой опасности. После чего результат такой модифицированной реакции оценивается и фиксируется уже для обобщенного профиля, который, тем самым, становится уже не новым и в последствии при его повторении не потребует вмешательства сознания, что может быть показано следующей схемой:
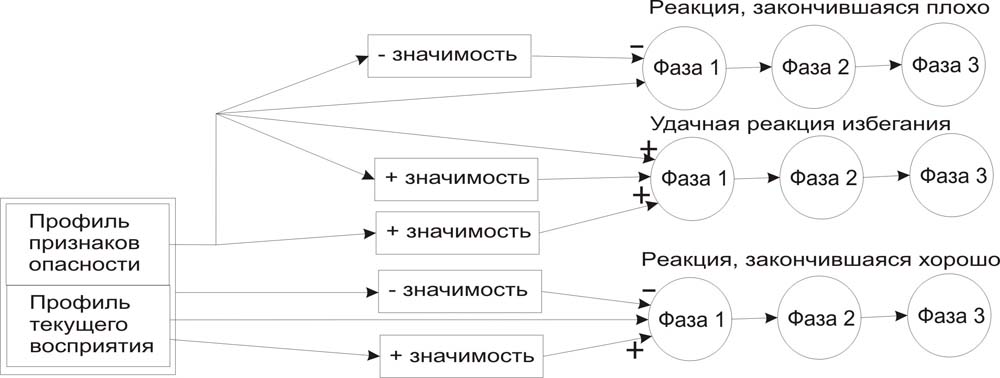
Этими схемами предположительно описаны самые примитивные пути модификации поведения в зависимости от условий. Достаточно древние механизмы используют предопределенности эмоциональных контекстов, которые оценивают результат реакции не просто как хорошо или плохо, а более специфично разделяют эти состояния, определяя преимущественный стиль поведения. Базовые (предопределенные) контексты формируются с помощью разделения распознавателей восприятия по видам медиаторов, а более молодые и более специфичные контексты - по выделением вниманием по признакам общих условий восприятия. Все это приводит к тому, что жизненный опыт (модификациолванное поведение) привязывается к стилю текущего поведения или условиям и формируется взаимно независимо так, что опыт в одном контексте оказывается автономным (и во многом недоступен) по отношения к другим контекстам.
Кроме того, эволюция механизмов сознания развила его функции до возможности экспериментировать с имеющимися цепочками памяти прежнего опыта без непосредственного реагирования так, что при этом легко прогнозируется результат различных возможных вариантов и даже имеется возможность синтеза новых вариантов на основе известных.
Сложность описания подобных функций и множество вариантов возможной реализации не позволяет их формализовать в этой статье увереннее, чем на уровне предварительных предположений. Но все они в полной мере используют все иерархии сложности нейронных сетей, которые были описаны. Используя понимание их организации, становится вполне возможным предположить возможные их реализации.
Целостные представления о функциях сознания в тезисной форме изложены в статье Сознание и эвристика.
В качестве наглядного обобщения текущего состояния понимания механизмов организации адаптивного поведения: Схема основных функциональных блоков в организации адаптивности поведения и их взаимосвязей.
Что хорошо в эволюционно-иерархическом подходе, так это - возможность не строить адаптивную систему сразу в полную сложность ее функциональности, а начать, например, с уровня досознательной оценки. При этом, конечно, единной сетью с общим ее обучением не обойдешься. Необходима многоуровневая система отдельных сетей, начиная с распознавателей примитивов, распознавателей образов всех рецепторных полей с выходом на эффекторные распознаватели фаз действий с последовательным обучением с простейших распознавателей к более сложным на основе более простых. Программная реализация такой системы не представляется реально возможной. Здесь необходимы аппаратная реализация по типу один распознаватель (нейрон с входными синапсами) - одно устройство. В этом пране Intell уже начала выпускать комплектующие.
Понимание сути субъективного, самоощущения своего Я, произвольности и взаимодействия души тела: Понимание произвольности.
В короткой статье – ясная формулировка того, что нужно для понимания сути сознания: Что такое сознание – просто и ясно.
Дополнительно:
