Сетчатка
Относится к разделу Биология
Строение сетчатки
Сетчатка [615] покрывает около 70% внутренней поверхности глазного яблока, анатомически представляя собой тонкую оболочку, прилежащую на всём своём протяжении с внутренней стороны к стекловидному телу, а с наружной — к сосудистой оболочке глазного яблока. Зрительная часть сетчатки имеет неоднородное слоистое строение, доступное для изучения лишь на микроскопическом уровне и состоит из 8 следующих вглубь глазного яблока слоёв. Иногда встречается нумерация из 10 слоев, за счет добавления двух разделительных слоев-мембран.
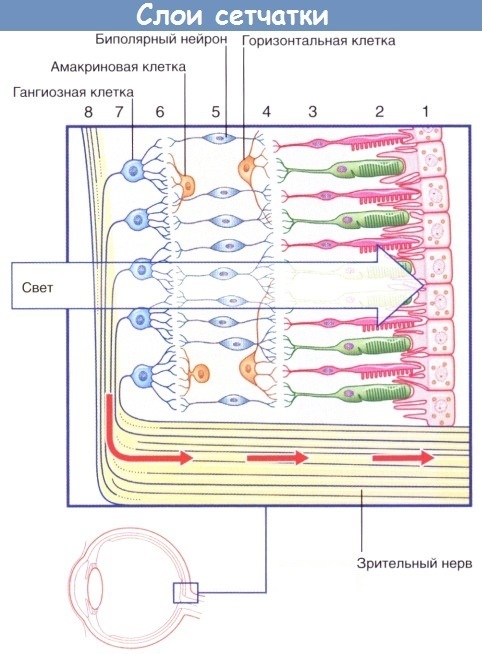
(1) Пигментный слой. (2) Фоторецепторный слой. (3) Наружный ядерный слой. (4) Наружный сетчатый слой. (5) Внутренний ядерный слой. (6) Внутренний сетчатый слой. (7) Слой ганглиозных клеток. (8) Слой нервных волокон.
По схеме видно, что сигнал с рецепторов (палочки и колбочки) поступает сначала на биполярные клетки, а с них на ганглиозные, которые передают его далее в мозг. При этом в схеме присутствуют еще горизонтальные и амакриновые клетки, выполняющие вспомогательную роль. Функциональное назначение клеток будет подробно рассмотрено ниже, пока лишь отметим для себя, что это совсем не похоже на фотоматрицу. Там схематически (но не в техническом исполнении) все достаточно просто:
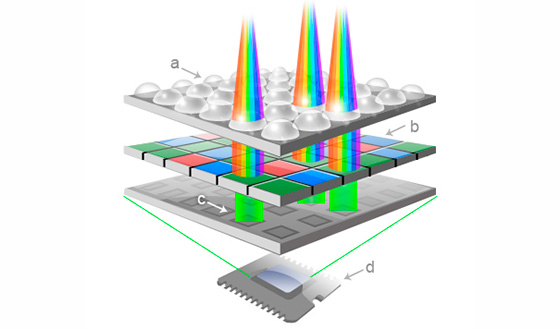
Через линзы свет падает на 3 типа фильтров (красный, синий, зеленый), равномерно распределенные по площади матрицы, которые лежат поверх рецепторов. В результате получаем возбуждение рецепторов, соответствующее составляющей цвета в данной точке-пикселе. Сам пиксель состоит из 4 ячеек, прикрытых фильтрами, причем зеленых там 2, потому, что человеческий глаз более восприимчив к нему.
Так устроен простейший фильтр Байера [635], а существующие более совершенные системы это по сути попытки создания матриц с максимально плотной упаковкой и различными типами кодирования цвета: RGGB, RGBW, RGEB, CGMY и т. д. Это не меняет сути того, что для получения кода цвета нужно использовать несколько базовых цветов. В графических программах обычно так и кодируют цвет, например: RGB(5,123,78), где цифры означают уровень яркости составляющей цвета, значение которых лежит от 0 до 255. В сетчатке используется аналогичный способ распознавания цветовых оттенков, но на этом подобие заканчивается и начинаются интересные варианты реализации, многие из которых до сих пор не поняты в полной мере.
Сетчатка относится к так называемым "инвертированным органам", расположена как бы "наизнанку": рецепторы в самом низу, на пигментном слое, нейроны, нервные волокна снаружи. Свет должен пройти через слой волокон зрительного нерва, слой ганглиозных клеток и слой биполярных нейронов, чтобы достичь фоторецепторов, поэтому они лишены миелина и прозрачны. Такое строение продиктовано в первую очередь необходимостью поглощать пигментным слоем рассеянный свет, не реагирующий с фоторецепторами, обеспечить их питанием и утилизировать их быстро распадающийся фотопигмент. То есть это просто особенность организации метаболизма данной биологической системы.
Фоторецепторы в сетчатке представлены двумя типами: палочки, максимально чувствительные к изменениям яркости (свету) и колбочки, способные различать длины волн (цвет). Они рассеяны по сетчатке не равномерно. В глазе человека расположено около 130 млн. фоторецепторных клеток, отношение палочек к колбочкам составляет 20:1 во всех отделах за исключением центральной ямки (фовеоле). Чувствительность палочек к свету и колбочек к цвету ограничена их физиологией, особенно колбочек, чей диапазон реагирования на длину волны относительно большой. Гиперчувствительность палочек к одиночному фотону и способность колбочек тонко различать оттенки цвета обеспечивается химическим каскадом усиления [626], взаимным перекрытием рецепторных полей, латеральным торможением.
Ближе к центру концентрация колбочек на единицу площади увеличивается и достигает максимума в центральной ямке, при этом уменьшается так же их размер. Поэтому такие колбочки называют карликовыми. Для максимальной плотности упаковки они размещены в ямке гексагональной (шестигранной) решеткой. На периферии концентрация колбочек уменьшается вплоть до полного отсутствия, при этом увеличивается количество палочек. В результате, в центральной ямке имеем область наиболее острого цветового зрения, которое становится менее острым и все более черно-белым по мере приближения к периферии.
Получается, что лишь центральную ямку, занимающую несколько процентов от общей площади сетчатки, можно назвать местом полноценного (острого) зрения, а остальная область выполняет вспомогательные функции детектирования. Думаю, основные причины неравномерного распределения в следующем:
- Эволюционно черно-белое зрение более древнее, так как более просто организовано, а все последующие находки эволюции строятся на базе предыдущих.
- При такой компоновке сетчатка представляет собой как бы несколько сенсорных систем, параллельно обрабатывающих зрительную информацию, что позволяет уже на уровне сетчатки получать ее в виде "срезов данных", отправляемых к своим специфическим отделам мозга, за счет чего формируется иерархическое комплексное реагирование: от простейших мгновенных реакций до сложно-анализируемых.
Сам принцип формирования все более усложняющегося абстрагирования подразумевает наличие иерархических систем реагирования, формирующих сначала простейшие рефлексы, а затем на их базе более сложные, где первичные служат фоновым контекстом для последующих и/или базовыми примитивами для формирования более сложных. Следовательно, стоит ожидать, что аналогичное по принципу строение имеют и прочие сенсорные системы (слуховая, вкусовая, обонятельная, осязательная).
Можно сказать, что сетчатка является многоканальным сенсорным органом, что принципиально отличает ее от фото-матрицы, которая просто регистрирует уровень активности цветных пикселов, равномерно распределенных по ее площади и передающих ее затем на АЦП (аналого-цифровой преобразователь) для получения массива данных снимка матрицы в двоичном коде.
Функциональность нервных клеток сетчатки
Большинство клеток нервной системы имеет химические синапсы. Такой нейрон возбуждаясь передает потенциал действия, что означает передачу аналогичного по силе возбуждения связанному с ним другому нейрону. Нервные клетки сетчатки, кроме ганглиозных, передают сигналы между собой электротонически, при помощи электрических синапсов [630]. Электрический синапс, судя по всему, эволюционно один из первых вариантов передачи сигналов между клетками, позволяющий развить активность нейронов максимально быстро, но из-за сопротивления мембран лишь на ограниченное расстояние. Такие синапсы обычно двунаправленного действия, что хорошо подходит для одновременной активации/дезактивации областей нейронов, выполняющих одинаковую функцию.
Одна из причин электротонического способа передачи сигналов это плотная упаковка нейронов в сетчатке и вероятно потребность максимально быстрой их передачи, для столь же быстрого реагирования. Это можно обосновать тем, что такие синапсы широко распространены у низших животных, где в основном требуются простейшие быстрые реакции. Для сетчатки это может быть, например, мгновенное реагирование на изменение освещенности, мелькание объекта в поле зрения. Но можно развить предположения дальше.
Исследования показали, что для палочек и колбочек величина их гиперполяризации прямо зависит от интенсивности освещенности. Передавая возбуждение через электрический синапс они передают не только потенциал действия, но и его величину. Судя по всему, это попытка реализации аналоговой передачи уровня сигнала с рецепторов на ганглионарную клетку через биполярные нейроны, хотя и в относительно небольшом диапазоне. Предположительно, это может выглядеть так: ганглионарная клетка, передающая активность сетчатки в мозг, связана с несколькими биполярными нейронами и ее активация зависит не только от количества связанных с ней активных биполяров, но и уровня сигнала, поступающего от них на ее электрические синапсы. В итоге она может активироваться несколькими биполярами, а может и одним, если он передаст достаточный по мощности сигнал. Получается интересная схема нейронных связей, в которой уровнем активности рецепторов можно менять схему подключения эффекторов, хотя физически схема не меняется. Как бы вариант ее модулирования, или даже правильнее сказать, конфигурирования входным сигналом. Но это лишь предположение, обоснованных утверждений, как еще нейроны сетчатки используют электрические синапсы мне не попадалось. С большей уверенностью можно предположить, что электротонический способ передачи возбуждения позволяет более тонко и плавно осуществлять контрастирование, без скачков переключения состояний нейронов. Так же возможно, что по этой же причине это менее энергозатратный вариант реагирования, когда вместо мощного разряда, требующего затем времени на регенерацию, нейрон лишь незначительно меняет уровень возбуждения. Это не только экономичная, но более динамичная (быстрая система) реагирования. Но как уже говорилось, у нее существенный недостаток, почти исключающий ее использование в более сложной нервной системе - сигнал быстро затухает.
Теперь рассмотрим более подробно цепочку реагирования нейронов сетчатки.
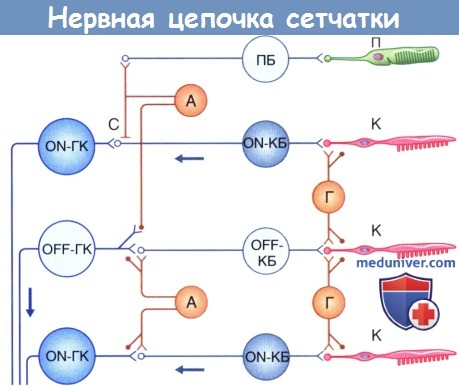
Схема нервной цепочки сетчатки: А—амакриновая клетка; К—колбочка; КБ—колбочковый биполярный нейрон; ГК—ганглиозная клетка; Г—горизонтальная клетка; С—соединение (щелевидный контакт); П—палочка; ПБ—палочковый биполярный нейрон.
Процесс возбуждения в сетчатке начинается с фоторецепторов [625], которые как уже говорилось, у животных с цветовым зрением представлены 2 типами клеток: свето-чувствительные палочки и цвето-чувствительные колбочки. Для цветового восприятия пришлось пожертвовать чувствительностью колбочек, так как специализация на диапазон длины волны означает пропорциональное уменьшение диапазона восприятия интенсивности, что обусловлено истощением рецепторов и необходимостью их регенерации. В итоге произошло разделение специализации: палочки детектируют интенсивность, а колбочки цвет.
Сигнал с фоторецепторов снимается при помощи биполярных нейронов [624], выполняющих роль связующих элементов между фоторецепторами и ганглиозными клетками, хотя исследования показывают, что некоторые из них способны самостоятельно детектировать уровень освещенности и предположительно служат детекторами движения. Как конкретно они это делают пока не ясно.
Биполярные нейроны имеют всего один аксон и один дендрит. Эти отростки отходят от противоположных концов клетки, которая обычно веретенообразной формы. Они соединяют через синапсы одну колбочку или несколько палочек зрительной системы с одной ганглионарной клеткой. Биполярные клетки палочек не образуют синапсов непосредственно с ганглионарными клетками, их синапсы находятся на амакриновых клетках.
Возникает вопрос, зачем потребовалось связывать ганглиозные клетки с рецепторами через биполяры, почему бы не напрямую? Думаю, дело в том, что это позволило создать схему получения сразу двух снимков активности рецепторов: прямую и инверсную, причем отдельно для палочек и колбочек. Для этого биполяры кроме специализации на палочковый и колбочковый, делятся еще на 2 типа: ON и OFF, которые активируются инверсно, что позволяет создать два потока взаимно-инверсных контрастирований, которые обрабатываются затем в зрительных колонках. Мне это напоминает маски в фотошопе для разных спектров.
Наличие специализированных ON/OFF нейронов говорит о том, что фиксируется не просто рецепторная активность палочек/колбочек, а изменения их активности. Это значит, что спроецированная на сетчатку статическая картинка регистрируется лишь на короткий промежуток времени и далее не воспринимается. Эксперименты показали, что если блокировать циркадные движения глаз при рассматривании статической картинки, она пропадает из поля зрения. Этот эффект эмпирически давно открыли мистики заметив, что при созерцании во время медитаций, в поле зрения остается лишь объект созерцания (пламя свечи, точка и т. п.). Такой алгоритм реагирования на раздражитель зрительных клеток сформировался по нескольким причинам:
- если как в фотоматрице рецепторы при раздражении все время будут активны, это быстро вызовет их истощение и возникнут серьезные проблемы с регенерацией.
- постоянный поток информации излишне перегружает каналы передачи, экономичнее для "трафика" передавать лишь измененния картинки, сохраняя в опреративной памяти предыдущий снимок
- разделение на ON/OFF реагирующие нейроны позволяет более эффективно регистрировать контрастные границы объектов
- специализация на изменении активности соседних рецепторов возможно позволяет создать алгоритм регистрации движения объекта с определением его направления
Исследования показали, что активность биполяров и ганглиозных клеток зависит не только от суммарного возбуждения их рецептивного поля фоторецепторов [631], но и от того, какая часть поля была более активна: центральная или периферия. Это обеспечивается при помощи связей горизонтальных и амакриновых клеток и позволяет реализовать два каскада контрастирования на уровне биполяров, а затем ганглиозных клеток. Мне видится в этом контрастировании попытка уменьшить сильно плавающий диапазон распознавания длины волны колбочек. За счет выделения активности центра, сопоставления с инверсной активностью периферии, перекрытия рецептивных полей в итоге получается сложная «интерферационная решетка цветовых контрастов», которая служит основой для определения оттенка в секторе рецепторной матрицы. К этому еще добавляется антагонистическое взаимодействие ганглиозных клеток, о чем будет ниже. И все эти сложности только потому, что нет в сетчатке аналога АЦП (аналого-цифрового преобразователя), а спектральный диапазон чувствительности колбочек слишком большой для точного детектирования цвета.
Смотрим схему дальше и видим, что между фоторецепторами и биполярами находятся горизонтальные клетки [618]. Выше уже говорилось, что они служат для организации взаимно-тормозного влияния между биполярными клетками, что обеспечивает эффект латерального торможения. Кроме этого они имеют обратную положительную и отрицательную связь с фоторецепторами, через так называемые положительные и отрицательные синапсы (обычно электрические синапсы двунаправленного действия), что еще больше усиливает эффект контрастирования. Мне кажется обоснованным сделать такой вывод: несмотря на большое разнообразие видов горизонтальных клеток у разных животных, их основное назначение - контрастирование активностей биполяров. Возможность управлять через латеральное торможение активностью целых областей биполяров означает по сути возможность получения конфигураций их «эффекторного рисунка». То есть на один раздражитель при разных уровнях торможения может быть сформирован свой уникальный ответный рисунок, а от него - рефлекторная цепочка. Это как я думаю, один из первичных, древних способов мгновенного реагирования. У простейших позвоночных как правило большая часть обработки зрительной информации происходит на уровне сетчатки, где и формируются конечные пусковые триггеры рефлексов. Подробнее о конфигурациях рассказывалось в статье ячеистая струткура нейросети, где на примере нескольких типов решеток показано, как меняются их эффекторные рисунки при изменении параметров связи элементов (уровень торможения, порог).
Фоторецепторы вместе с горизонтальными клетками и биполярами образуют так называемую триаду. Триада состоит из пресинаптической мембраны фоторецептора, в которую заходит дендрит биполяра, а по бокам от него заходят отростки горизонтальных клеток. Значение и принципы функционирования триады пока остаются неизвестными. Но предполагается, что за счет таких связей между фоторецепторами, горизонтальными клетками и биполярами, центр рецептивного поля биполяров формируется прямым путём при передаче сигнала от фоторецепторов, а периферия формируется за счет непрямого пути через горизонтальные клетки за счёт возбуждающих или тормозных электрических синапсов.
Следующими по схеме идут амакриновые клетки [619], передающие сигналы в двух направлениях: либо прямо от биполярных клеток к ганглиозным, либо горизонтально в пределах внутреннего сетчатого слоя от аксонов биполярных клеток к дендритам ганглиозных клеток или к другим амакриновым клеткам. Амакриновые клетки проводят сигналы латерально во внутреннем сетчатом слое, то есть являются аналогом тормозных горизонтальных клеток, но уже для ганглиозных, контрастируя их активность. Получается уже как бы второй каскад контрастирования, о котором уже говорилось выше.
В настоящее время морфологическими или гистохимическими методами идентифицированы около 30 типов амакриновых клеток. Функции некоторых из них охарактеризованы, и все они отличаются друг от друга. Один тип амакриновых клеток является частью прямого пути палочкового зрения, т.е. от палочки к биполярным клеткам, затем к амакриновым и, наконец, к ганглиозным клеткам. Амакриновые клетки другого типа активно реагируют в начале непрерывного зрительного сигнала, но ответ быстро исчезает. Некоторые амакриновые клетки, наоборот, мощно реагируют на выключение зрительного сигнала, но их ответ также быстро прекращается. Амакриновые клетки еще одного типа реагируют и на включение, и на выключение света, просто сигнализируя об изменении освещения, независимо от его направления. Есть амакриновые клетки, реагирующие на движение пятна света по сетчатке в определенном направлении; о таких клетках говорят, что они чувствительны к направлению. В известном смысле большинство амакриновых клеток являются вставочными нейронами, которые помогают анализировать зрительные сигналы, прежде чем они покинут сетчатку.
Складывается впечатление, что амакриновые клетки являются частью более древнего черно-белого зрения, с вариантами распознавания простейших фоновых возбуждений типа: что то мелькнуло, внезапно потемнело/посветлело и т.д. Видимо особо не заморачиваясь схемами связей, природа просто выдумала множество специфических типов клеток, которые и осуществляют подобное детектирование. Хотя и не совсем понятно, как именно можно определить направление движения пятна света по сетчатке, но как показывают исследование, как то это все же определяется, о чем говорит детектирование активности соответствующей клетки на свой тип раздражителя. Но при реализации искусственной сетчатки думаю подобные извороты можно переложить на специальные функции, потому, что на сегодняшний начальный уровень развития нанотехнологий проще придумать алгоритм распознавания, чем специфический рецептор. А здесь важно лишь снова отметить, что исследования однозначно подтверждают сложность, я бы сказал «многоканальность» выходного сигнала сетчатки. И это косвенно подтверждает предлагаемый принцип построения иерархии реагирования в нервной системе: от простой реакции к сложной, с передачей ей управления дальнейшим ветвлением рефлекторной цепочки.
Конечным звеном нейронной активности сетчатки являются ганглиозные клетки [620], проводящие сигналы через зрительный нерв за пределы сетчатки в ствол мозга. Единственные нейроны сетчатки, сигналы которых всегда передаются в виде потенциалов действия, идущих по зрительному нерву к мозгу.
Как уже говорилось выше, по мере приближения к центральной ямке все меньше палочек и колбочек конвергируют на каждое зрительное волокно. Кроме того, и палочки, и колбочки становятся тоньше. Эти эффекты постепенно увеличивают остроту зрения в центральной сетчатке. В самом центре есть только тонкие колбочки (примерно 35 000) и совсем нет палочек. К тому же, количество волокон зрительного нерва, выходящих из этой части сетчатки, почти равно числу колбочек. Это объясняет высокую степень остроты зрения в центральной сетчатке по сравнению с гораздо меньшей остротой на периферии.
Другим различием между периферией и центром сетчатки является гораздо более высокая чувствительность периферической сетчатки к слабому свету. Отчасти это объясняется тем, что чувствительность палочек в 30-300 раз выше, чем колбочек. Однако эффект значительно усиливается в связи с конвергенцией примерно 200 палочек на одну ганглиозную клетку в периферических областях сетчатки, поэтому сигналы от палочек суммируются, дополнительно усиливая интенсивность стимуляции периферических ганглиозных клеток и исходящих от них волокон зрительных нервов. Различают три группы ганглиозных клеток, которые обозначают как W-, Х- и Y-клетки. Каждая группа выполняет свою функцию.
- W-клетки – передают сигналы с низкой скоростью, около 8 м/с, мелкого размера. Их около 40% от общего количества. В основном возбуждаются от палочек (через биполяры и амакриновые клетки). Их рецептивные поля широкие и потому предполагается, что они чувствительны в первую очередь к регистрации направленного движения и для грубого палочкового зрения в темноте
- Х-клетки – большая часть (55%) ганглиозных клеток, скорость передачи сигнала средняя (14 м/с). Их рецептивные поля не большие, включают в охват минимум одну колбочку. Из этого делается предположение, что они служат для детализации изображения и цветопередачи.
- Y-клетки – самые крупные ганглиозные клетки, проводящие сигнал с большой скоростью 50 м/с и выше. Доля их всего 5% от общего количества, но зато они имеют широкие рецептивные поля, поэтому предполагается, что они служат для быстрой регистрации изменений (движений или освещенности) в поле зрения, но без высокой точности. Это может служить например для синхронизации циркадных движений глаз или других рефлекторных реакций, требующих быстроты реагирования.
Отличие ганглиозных от остальных клеток сетчатки в том, что даже при отсутствии стимуляции они посылают постоянные импульсы с частотой 5-40 в секунду. Зрительные сигналы, в свою очередь, накладываются на эту фоновую активность ганглиозных клеток. Тут как бы напрашивается аналогия с амплитудной модуляцией, но одновременно встает вопрос – зачем это надо? Но это не все сюрпризы ганглиозных клеток.
Долгое время считалось, что палочки и колбочки являются единственными фоторецепторами сетчатки, пока в 1991 году не открыли светочувствительные ганглиозные. Они не участвуют в формировании зрительного сигнала, но способны различать уровень освещения, что позволяет им принимать участие в регулировании циркадных ритмов у птиц и млекопитающих, а так же ширины зрачка в зависимости от освещения.
Распознавание оттенка [623] цвета начинается в слое ганглиозных клеток, где некоторые из них возбуждаются колбочкой одного цветового типа и тормозятся колбочкой другого цветового типа. Например, красные колбочки вызывают возбуждение специфической ганглиозной клетки, а зеленые ее торможение, или наоборот. Такие связи называют реципрокными. Аналогичный антагонизм существует между синими и сочетанием красных и зеленых колбочек. Таким образом, каждый цветоконтрастный тип ганглиозной клетки возбуждается одним цветом, но тормозится «оппонентным» цветом. Это позволяет существенно улучшить распознавание оттенка при перекрытии рецептивных полей [631] ганглиозных клеток, с учетом того, что диапазон реагирования колбочек довольно большой и соответственно точность цветовосприятия низкая.
Тут нужно сделать одно уточнение по поводу цветовосприятия, которое обычно не замечается - способность воспринимать цвета в независимости от цвета освещения. В самом деле, мы воспринимаем в основном отраженный от предметов свет на фоне многократно отраженного света от других предметов, и если бы не была развита способность выделять базовый цвет из смеси всех прочих - была бы постоянная путаница. Такую способность называют контрастным восприятием окраски, и судя по назаванию здесь так же не обходится без латерального торможения (контрастирования) близлежащих рецептивных полей. Можно предположить, что вышеназыванный цветовой антагонизм как раз и возник как приспособление для выделения базового цвета объекта. Конечно при достаточно сильном фоновом, и тем более монохромном освещении возникают искажения цветовосприятия, но это уже искуственнно освещение.
Теперь можно подвести итоги:
Очевидно, что сетчатка лишь отдаленно напоминает фотоматрицу, в основном тем, что в ней так же используется определение цвета через триаду базовых цветов: красный, синий, зеленый. Но в отличие от матрицы в сетчатке происходит первичная обработка сигналов через контрастирование активностей связанных с рецепторами нервных клеток и детектирование простых примитивов специфическими рецепторами, реагирующими на повышение/понижение активности внешнего раздражителя (ON/OFF ответ рецепторов). Различные по размеру, от одиночных рецепторов до обширных областей, перекрывающиеся рецептивные поля нейронов сетчатки формируют сдвинутые по фазе, разные по характиристикам и длительности сигналы, передающиеся по отдельным связям. Электротонический способ передачи возбуждения в сетчатке используется по причине плотной упаковки нейронов, а так же предположительно как менее энергозатратный, более быстрый и позволяющий точнее регулировать контрастирование активностей. В результате, выходной сигнал разделяется на отдельные потоки, которые с разными скоростями направляются в соответствующие отделы зрительной коры, причем поток двунаправленный - происходит подстройка на лету параметров для уточнения образа. Это позволяет формировать иерархические системы реагирования на разных уровнях: от простейших рефлексов до сложных распознаваний образов. То есть формирование и передача примитивов восприятия в зрительную кору судя по всему идет по схеме:
- изменился фон освещения - возникшая реакция становится контекстом для формирования последующих примитивов
- что то мелькнуло - определяется примерная скорость и направление, активируются следящие за объектом системы, происходит захват изображения. В зависимости от полученных предварительных параметров добавляется новый контекст (мелькнуло быстро/медленно, сверху/снизу/в сторону и т. д.)
- включается сканирование захваченного в поле зрения объекта, происходит первичное разбиение на простейшие примитивы, группировка признаков и добавление нового контекста, все более сужающего область поиска подходящей реакции (определилось что то прямоугольное, круглое, черно-белое, цветное и т. д.)
- после предварительной обработки полученных примитивов включается более точное сканирование выбранных областей объекта, для окончательноно определения что это и выбора подходящей реакции.
Если бы сетчатка была устроена по аналогии с фотоматрицей, где просто считываются данные рецепторов и после преразования через АЦП передаются на программную обработку - было бы не возможно организовать иерархическое формирование реакций без привлечения сложного мат. аппарата, который вряд ли возможно создать на элементной базе в виде нейронов методом естественного отбора. Но благодаря сложной предварительной обработке активностей нервных клеток сетчатки итоговая информация представляет уже практически сформированные примитивы - контуры, которые остается только сопоставить, что на порядок проще. Поэтому усложнение зрительной системы всегда происходило как параллельное усложнение рецепторных органов (сетчатка) и принимающих отделов (зрительная кора).
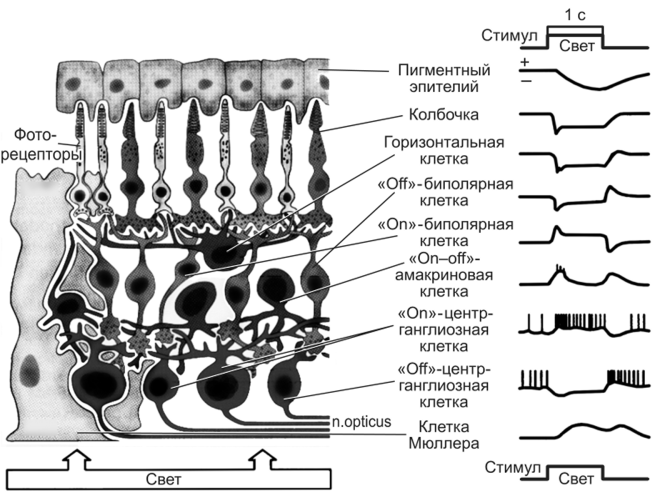
Виды электрических потенциалов (справа), генерируемых клетками различных слоев сетчатки при действии света (R.F. Schmidt, F. Lang, G. Thews, 2005)
Это важно понимать, чтобы не поддаваться на спекуляции типа "голографической памяти", "волновой теории сознания", "интеллекта насекомых" и т. п., когда пытаются понять работу нервной системы мало обращая внимания на устройство рецепторных органов, хотя именно там происходит первичная обработка информации, часто очень сложная, и это снимает многие вопросы типа "как мозгу удается понять...". На самом деле часто это уже удалось сделать на уровне рецептора. Это особенно заметно на примере насекомых, где например по структуре фасеточного глаза даже визуально заметна его функциональная неоднородность: у летунов обычно верхние фасеты настроены на восприятие поляризации, нижние на цвет и форму объектов. У хищников типа богомола критически важна детализация и глубиномер, поэтому по центру глаза у них расположена фовеа с максимально плотной упаковкой фасет. Глаза насекомых часто сверкают словно радуга, переливаясь сложными интерференционными рисунками, показывая специализацию отдельных фрагментов глаза для выполнения специфических задач - посылая в итоге многоканальный отформатированный сигнал в зрительные ганглии, существенно упрощая задачу анализа информации.
Если задуматься о создании искусственного аналога сетчатки, подходящим для систем с ИИ, то возникают следующие соображения:
- Искусственный прототип так же должен выдавать несколько потоков сигналов разного уровня абстрагирования, иначе не получится сформировать иерархическую систему реагирования. Это принципиально.
- Для реализации многопоточной обработки можно пойти природным путем, разделив рецепторы на специальные типы и разместив их неравномерно по площади матрицы и добавив в схему связи вспомогательные компоненты, аналоги биполяров, горизонтальных, ганглиозных и амакриновых клеток. Это потребует существенно более продвинутого развития био-инженерных технологий. Можно пойти программным, добавив после АЦП дополнительный модуль форматирования и разделения на потоки сигналов. Но думается, это потребует очень серьезных вычислительных мощностей и более целесообразно сделать смешанный вариант: переложить часть функционирование на элементы, а часть на программную обработку. Например, как показали попытки моделирования фрагментов нейросети на конструкторе нейросхем есть принципиально не решаемая проблема расчета состояний элементов, обусловленная тем, что современные компьютеры работают в псевдо-параллельном режиме вычислений, а требуется действительно параллельное, которое не возможно осуществить на ЭВМ с централизованным управлением одним процессором. В нервной система однако все процессы протекают параллельно. В итоге симуляция на конструкторе вынужденно приводит к созданию фонового бесконечного цикла расчета, что сильно его затормаживает и в настоящее время не может быть и речи о симуляции связей миллионов нейронов, не говоря уже и миллиардах. Но если часть взаимодействий элементов искусственной сетчатки реализовать в виде специфических компонентов, связанных аналогичным образом как в природной реализации, мы получим полноценный прототип и существенно разгрузим процессор, заняв его например реализацией уж слишком специфических распознавателей. То есть мне пока что однозначно видится практическая реализация именно как смешанный вариант.
- Мне не раз попадались рассуждения от профильных специалистов, занимающихся разработкой искусственных нейросетей, что не смотря на очевидные успехи в плане повышения скорости и качества распознавания, так же повышается уверенность движения не в ту сторону. Потому, что нет понимания, что со всем этим растущим качеством делать дальше, как превратить его в полноценный «интеллект», а не рефлекторное реагирование, чем он пока что является. Мне же интересно другое: можно ли что то использовать из огромного числа алгоритмических наработок, созданных в этой области при создании прототипа природных интеллектуальных сенсорных систем или просто не обращать внимания, раз они сами говорят, что идут не в ту сторону? Думаю, ответ так же где то по середине. Природа была ограничена в компонентах и тем более в алгоритмах, все эволюционные находки строились на базе предыдущих, а это накладывает определенные ограничения и создает проблемы. Самый наглядный пример – операционные системы, вынужденные долго поддерживать устаревшие наработки единственно потому, что иначе придется переписывать множество программ, что вызовет большие проблемы. Поэтому нет смысла слепо копировать за природой ее находки, которые обычно являются заплатками.
Список произведений >>
Список публикаций >>
Тест: А не зомбируют ли меня? Тест: Определение веса ненаучности
| Обнаружен организм с крупнейшим геномом Новокаледонский вид вилочного папоротника Tmesipteris oblanceolata, произрастающий в Новой Каледонии, имеет геном размером 160,45 гигапары, что более чем в 50 раз превышает размер генома человека. | Тематическая статья: Формирование субъективного опыта в онтогенезе |
Рецензия: Работы А.Иваницкого, комментарии | Топик ТК: Свойства осознанного внимания и циклы |