
В мозге птиц обнаружен механизм, не позволяющий переучивать выученное
Системная нейрофизиология (nan)
Актуальная значимая новизна формирует новый вариант реагирования.
Вес уверенности: Вполне уверенно подтверждается независимыми исследователями
 Рис. 1. Взрослый самец зебровой амадины учит молодого самца правильно петь. Кадр из видеофильма, прилагающегося к обсуждаемой статье в Science |
Американские нейробиологи обнаружили механизм, не позволяющий зебровым амадинам переучивать те элементы песни, которые они уже выучили. Ключевую роль в обучении пению играют премоторные нейроны верхнего вокального центра (HVC), которые активно работают у молодых самцов («учеников») при прослушивании песни взрослого самца («учителя»). У взрослых самцов эти нейроны перестают реагировать на звуки чужих песен. Однако, как выяснилось, дело тут не столько в возрасте, сколько в уровне вокального мастерства ученика и в активном торможении работы премоторных нейронов HVC другими нейронами того же отдела мозга. Вставочные нейроны HVC посылают тормозящие сигналы премоторным нейронам при проигрывании тех элементов песни учителя, которые ученик уже умеет хорошо воспроизводить, и не делают этого, если звучит еще не освоенная учеником трель. Если у млекопитающих тоже есть подобные механизмы, то данное открытие, возможно, позволит в будущем научиться преодолевать косность мышления.
Зебровые амадины — удобный объект для исследования нейрологических механизмов обучения. Самцы этих птиц в молодости активно учатся петь, слушая пение старших (рис. 1). Ученым уже удалось выявить ключевые отделы мозга, задействованные в этом процессе. Показано, что для успешного обучения необходима работа премоторных нейронов верхнего вокального центра (HVC). Эти нейроны получают сигналы от слуховых отделов мозга и посылают нервные импульсы в ядро аркопаллиума (Robust nucleus of arcopallium, RA); рис. 2.
< Блок иллюстрации >

В опытах с птицами, находящимися под наркозом, было показано, что премоторные нейроны HVC самцов реагируют генерацией потенциалов действия на пение других самцов. Если заблокировать работу этих нейронов, молодой самец не научится петь.
В новом исследовании, результаты которого опубликованы в свежем выпуске журнала Science, американские нейробиологи раскрыли неизвестные ранее детали процесса обучения. На этот раз опыты проводились с бодрствующими самцами, которых аккуратно фиксировали, чтобы они не шевелились во время эксперимента. Через крошечные отверстия в черепе птицам вживляли микроэлектроды в премоторные нейроны HVC, аксоны которых идут в RA. Это позволило следить, во-первых, за активностью самих этих нейронов (регистрировались генерируемые ими потенциалы действия, то есть «выходящие» сигналы, поступающие из HVC в RA), во-вторых, за получаемыми ими «входными» сигналами от других нейронов. Такие сигналы бывают двух типов: возбуждающие и тормозящие. Они приводят к открытию тех или иных ионных каналов в мембране нейрона и к возникновению электрических токов (потоков ионов), которые стремятся либо ослабить, либо усилить поляризацию мембраны, что, в свою очередь, либо повышает, либо снижает вероятность того, что нейрон сгенерирует потенциал действия.
Для начала авторы посмотрели, как реагируют премоторные нейроны HVC самцов разного возраста на песню взрослого самца. Оказалось, что у молодых слушателей эти нейроны реагируют на звук песни, а у взрослых — нет. При этом у молодых самцов отдельные нейроны при повторных прослушиваниях раз за разом «выстреливали» в одни и те же, строго определенные моменты, соответствующие тем или иным элементам песни. Нейроны RA, получившие сигналы от HVC, тоже реагировали на это генерацией нервных импульсов. По-видимому, эта активность и лежит в основе обучения.
Теперь нужно было выяснить, почему у взрослых самцов премоторные нейроны HVC не реагируют на звук чужой песни. Может быть, они перестают получать входные сигналы от слуховых центров? Однако это предположение не подтвердилось: измерения возбуждающих постсинаптических токов (excitatory currents) показали, что возбуждающие сигналы при звуках песни исправно приходят на премоторные нейроны HVC у всех самцов независимо от возраста.
Почему же у взрослых самцов нейроны перестают реагировать на эти сигналы? Логично предположить, что тут замешано активное торможение. В пользу этого свидетельствовал и тот факт, на первый взгляд противоречащий обсуждаемым результатам, что ранее в опытах на птицах, находящихся под наркозом, было показано, что звук песни вызывает реакцию премоторных нейронов HVC и у взрослых самцов тоже. Авторы обратили внимание, что анестетик, использовавшийся для усыпления птиц в этих опытах, — уретан (см. Ethyl carbamate) — препятствует передаче тормозящих сигналов в мозге, блокируя работу важнейшего тормозного нейромедиатора ГАМК.
Чтобы проверить гипотезу о торможении, авторы ввели в HVC взрослых самцов вещество, блокирующее рецепторы ГАМК, — габазин — и проверили, как теперь будут реагировать премоторные нейроны HVC на звук песни. Предположение подтвердилось: после этой процедуры нейроны у взрослых самцов стали реагировать на песню так же, как у молодых.
Таким образом, реакция премоторных нейронов HVC на звук песни (и, следовательно, процесс обучения) зависит не от возбуждающих сигналов, приходящих от слуховых центров (эти сигналы приходят в любом случае), а от наличия или отсутствия тормозящих сигналов.
Дальнейшие эксперименты показали, что источником этих тормозящих сигналов являются вставочные нейроны того же отдела мозга (HVC). Правда, общая картина их реакции на звук песни оказалась сложной и запутанной, и с возрастом птицы она не связана (по крайней мере, не связана напрямую, каким-то простым и очевидным образом).
Но авторам все-таки удалось обнаружить в этом хаосе важную закономерность. Они рассудили, что смысл торможения может состоять в том, чтобы прекращать обучение, когда самец уже научился петь достаточно хорошо. В конце концов, нельзя же вечно перестраивать свои нейронные контуры и переучиваться всякий раз, заслышав песню нового соседа. Если навык уже приобретен и доведен до приемлемого уровня эффективности, то его следует зафиксировать и больше не трогать. Может быть, это сужает горизонты и делает ваше мышление несколько косным, зато гарантирует сохранение усвоенных знаний.
Чтобы проверить это предположение, авторы сопоставили работу вставочных нейронов HVC при звуке песни с певческим мастерством слушателя. Гипотеза блестяще подтвердилась. Оказалось, что интенсивность реакции вставочных нейронов HVC на звук песни, а также степень согласованности импульсов с определенными элементами песни тем выше, чем лучше данный самец воспроизводит песню учителя.
Таким образом, если самец уже научился петь в точности как учитель, то вставочные нейроны HVC при звуках этой песни посылают премоторным нейронам тормозящие сигналы, и поэтому те не реагируют на звук, хотя по-прежнему получают возбуждающие сигналы от слуховых отделов мозга. В результате самцы, уже научившиеся хорошо петь, больше этому искусству не учатся.
Песня самца зебровой амадины состоит из нескольких различающихся элементов — «слогов». При этом самцы не учат всю песню целиком как нечто единое и неделимое, а зазубривают слоги по очереди, подолгу тренируясь и отшлифовывая каждый слог. Поскольку было замечено, что реакция многих нейронов HVC (и премоторных, и вставочных) приурочена к строго определенным местам прослушиваемой песни, авторы предположили, что торможение может быть выборочным: уже выученные трели стимулируют активность вставочных нейронов, а еще не освоенные — подавляют ее.
Чтобы это проверить, были изготовлены записи двух искусственных песен, составленных из одного или двух слогов (их условно назвали А и B), вырезанных из настоящей песни. Первая искусственная песня имела вид АААА, вторая — ABAB. Необученным молодым самцам сначала давали слушать первую песню. В результате они обучались воспроизводить слог А, а слог B оставался для них незнакомым. Потом им проигрывали вторую песню, регистрируя работу нейронов во время прослушивания. Результаты показаны на рис. 3.
< Блок иллюстрации >
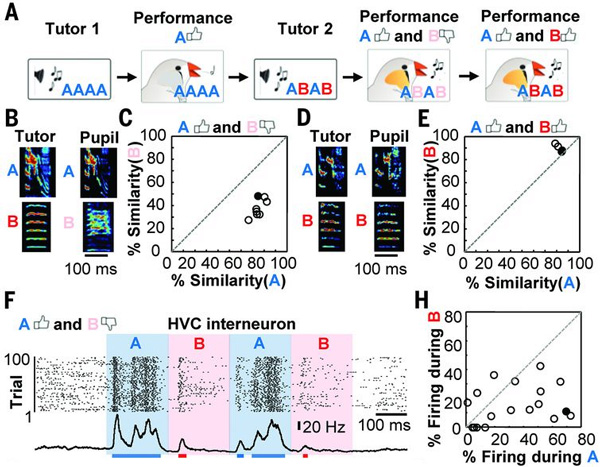 Рис. 3. Результаты эксперимента с искусственными песнями из двух слогов. A — схема обучения. Сначала необученному самцу много раз прокручивают песню, состоящую только из слога А (Tutor 1). В результате ученик выучивает этот слог. Затем начинается обучение при помощи второй песни, состоящей из чередующихся слогов A и B (Tutor 2). Сначала ученики хорошо воспроизводят слог A и плохо — слог B, а затем начинают хорошо воспроизводить оба слога. На разных этапах обучения регистрировалась работа нейронов при прослушивании песни. B — фонограммы исполнения слогов A и B учителем (Tutor) и учеником (Pupil) на первом этапе обучения, когда первый слог уже выучен, а второй еще нет. C — график, отражающий сходство слогов A и B в исполнении учителя и ученика на первом этапе обучения. D, E — то же самое для второго этапа обучения, когда ученик уже умеет хорошо исполнять оба слога. F — активность вставочных нейронов HVC в мозге ученика, находящегося на первом этапе обучения, во время прослушивания песни «ABAB». Видно, что вставочные нейроны резко усиливают свою работу, когда звучит уже выученный учеником слог А. H — соотношение активности вставочных нейронов при звучании слогов A и B. Черный кружок соответствует нейрону, работа которого показана на рисунке F. Изображение из обсуждаемой статьи в Science |
Как и следовало ожидать, во время звучания уже освоенного слога А активность вставочных нейронов HVC резко усиливалась, а работа премоторных нейронов, соответственно, тормозилась. Когда же звучал незнакомый слог B, наблюдалась обратная картина: активность вставочных нейронов снижалась, премоторных — усиливалась. Когда же ученики выучивали слог B, активность нейронов при звучании обоих слогов становилась одинаковой.
Таким образом, вставочные нейроны HVC откуда-то знают (откуда — пока неизвестно), какие элементы песни уже выучены, а какие нет, и при звучании выученных слогов генерируют импульсы, тормозящие активность премоторных нейронов HVC. В результате процесс обучения блокируется. Если же звучит еще не выученный слог, вставочные нейроны снижают свою активность, и тогда премоторные нейроны реагируют на звук генерацией нервных импульсов, которые передаются в ядро аркопаллиума (RA), и в результате самец выучивает новый слог.
Конечно, нерешенных вопросов осталось очень много. Неизвестно, откуда вставочные нейроны получают информацию о том, выучен данный слог или нет. Неизвестно, сколько разных слогов должен выучить самец и насколько сложной должна стать его песня, чтобы обучение новым слогам прекратилось. Неизвестно, задействованы ли нейроны HVC и RA, участвующие в обучении пению, также и в самом пении (это считается вероятным по аналогии с зеркальными нейронами премоторных и моторных областей коры млекопитающих, но не доказано). Неизвестно, насколько широко распространен в животном мире обнаруженный у зебровых амадин механизм торможения нейронов, ответственных за обучение. Кроме того, у взрослых бодрствующих самцов премоторные нейроны HVC перестают реагировать на звуки чужой песни даже в том случае, если слушатель петь так и не научился, и почему это происходит — тоже неизвестно.
Обилие вопросов, оставшихся без ответа, не умаляет значения обсуждаемой работы. Что-то подсказывает, что найденный авторами удивительный механизм «выучивания раз и навсегда», сопряженный с активным противодействием попыткам переучивания, есть не только у амадин. И тогда можно надеяться, что открытие этого механизма и его дальнейшее изучение позволит нам когда-нибудь научиться регулировать косность собственного мышления. У амадин, как мы видели, сделать это очень просто: нужно всего лишь ввести в строго определенную точку мозга немного габазина. Хотя, конечно, побочные эффекты этой процедуры никто пока не изучал.
Источник: Daniela Vallentin, Georg Kosche, Dina Lipkind, Michael A. Long. Inhibition protects acquired song segments during vocal learning in zebra finches // Science. 2016. V. 351. P. 267–271.
Источник:
>>Если навык уже приобретен и доведен до приемлемого уровня эффективности, то его следует зафиксировать и больше не трогать. Может быть, это сужает горизонты и делает ваше мышление несколько косным, зато гарантирует сохранение усвоенных знаний.... если самец уже научился петь в точности как учитель , то вставочные нейроны HVC при звуках этой песни посылают премоторным нейронам тормозящие сигналы, и поэтому те не реагируют на звук. Описан типичный механизм определения новизны, но не любой, а значимой - для задачи культурной преемственности. Всякие другие звуковых последовательности не запоминаются.
>> у взрослых бодрствующих самцов премоторные нейроны HVC перестают реагировать на звуки чужой песни даже в том случае, если слушатель петь так и не научился Новизна есть, но актуальной значимости освоить важную для птичьей коммуникабельности песню уже нет. Значимость здесь - потерянная исследователями компонента.
Проблема этих исследователей - непонимание общих принципов адаптивности и поэтому невозможность правильно, более системно планировать эксперимент.
Последнее редактирование: 19.01.2016
Относится к аксиоматике: Системная нейрофизиология.
Другие страницы раздела "Новизна-значимость":
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя