
ЗЕРКАЛЬНЫЕ НЕЙРОНЫ, ФИЗИОЛОГИЧЕСКАЯ РОЛЬ, ОСОБЕННОСТИ ФУНКЦИОНИРОВАНИЯ И ЭМОЦИОНАЛЬНО НАСЫЩЕННАЯ КОГНИТИВНАЯ КАРТА МОЗГА
Системная нейрофизиология (nan)
Зеркальным нейронам приписываются функции, которые относятся к субъективным ощущениям, в том числе и процессы самосознание человека.
Зеркальных клеток очень мало (в нижней теменной доле двух макак – резусов, была зафиксирована активность всего 41 зеркального нейрона), они разбросаны по коре мозга маленькими островками, но их функции вовлекают большие нейронные сети. Проблему можно решить, если предположить, что зеркальные нейроны функционируют как нейроны “бабушки”. В этом случае функции, которые приписываются зеркальных нейронов, превращаются в функции коры больших полушарий, которая является частью эмоционально насыщенной когнитивной карты мозга.
Процессы осознания и идентификации – это взаимодействия “Down Up” сигнала с “Top Down” сигналом из префронтальной коры. Эмоционально насыщенная когнитивная карта мозга формируется интеграцией нейронов гиппокампа нейронами коры больших полушарий. Наше “Я”, через процессы само-осознание, само-идентификацию и виртуальный скачок взаимодействуют с нейронами больших полушарий, получающих информацию о процессах в окружающей среде и формирующих контекст окружающей среды, представляющий собой наше субъективное ощущение. Новая информация о процессах в окружающей среде, консолидируется эксплицитной памятью с помощью специфической модификации экспрессии генов, и хранится в открытом доступе, то есть в состоянии быстрого воспроизведения при напоминании.
В общей сложности 15 зеркальных нейронов энергично сработали, когда обезьяна наблюдала движение “схватить, чтобы съесть”, но не обнаруживали никакой активности, пока экспериментатор хватал яблоко и просто держал в руке. Других 4 зеркальных нейрона активировались в ответ на то, что экспериментатор в конце концов помещал яблоко в чашку, но не съел его. Только тип действия, а не кинематическая сила, с которой моделируются объекты, определяли активность нейронов.
Также было важно, что нейроны срабатывали до того, как обезьяна видела поведение человека, запускающего второй моторный акт – поднесение объекта ко рту или помещение его в чашку. Следовательно, нейроны нижней теменной доли кодируют одно и то же действие (хватание) по-разному В СООТВЕТСТВИИ С КОНЕЧНОЙ ЦЕЛЬЮ ДЕЙСТВИЯ. Они могут предоставить нейронную основу для прогнозирования последующих действий другого человека и определения намерения.
Другая возможная функция зеркальных нейронов – это облегчение обучения. Зеркальные нейроны кодируют (не реально, а виртуально) конкретное представление действия, то есть, представление, которое активировалось бы, если бы наблюдатель действовал. Это позволило бы нам моделировать (внутренне повторять) наблюдаемое действие неявно (виртуально в мозге), чтобы быстрее активировать наши собственные двигательные программы наблюдаемых действий и подготовиться к воспроизведению действия, через некоторое время. Это неявное обучение. Благодаря этому, наблюдатель будет реализовать свое действие с большей ловкостью.
Обучение с помощью зеркальных нейронов лучше назвать “Обучение через подражание или обучение с учителем”. Когда родители обучают детенышей охотиться, то детеныши наблюдают как это делают родители и подражая им сами обучаются охотиться.
Зеркальные нейроны кодируют целенаправленные действия, потому что они активируются, если наблюдаемое действие является целенаправленным. Однако зеркальные нейроны активируются только тогда, когда наблюдаемое действие направлено на цель (объектно-ориентированное действие или коммуникативный жест, который, безусловно, также имеет цель). Как они “узнают”, что определенное действие направлено на достижение цели? На каком этапе их активации они обнаруживают цель движения или его отсутствие? По его мнению, зеркальная нейронная система может быть активирована только после того, как цель наблюдаемого действия прописывается некоторыми другими структурами мозга.
Возможно, человеческий язык развился из системы представления/понимания жестов, реализованной в зеркальных нейронах. Показатели расширения словарного запаса связаны с возможностями детей вокально имитировать не слова, а похожие звуки и, таким образом, приобретать новые произношения слов. Такое повторение речи происходит автоматически, быстро и облегчает восприятие речи. Кроме того, такая вокальная имитация может происходить без понимания.
Прочное представление времени и пространства в гиппокампе предполагает фундаментальный механизм для формирования пространственно-временного континииума в рамках согласованных воспоминаний. Можно полагать, что клетки времени, совпадающие по особенностям функционирования с клетками места, выстраивают внешние события во временной последовательности, так как активность клеток места меняется и при изменении внешних событий. Это очень важно, так как память консолидирует временную последовательность событий, и память воспроизводится во временной последовательности.
Целенаправленное поведение это активный поиск объектов удовлетворения потребности, которая контролируется когнитивной картой и подкрепляется или не подкрепляется оценочной функцией эмоциональных состояний. Оценочная функция, точнее формирование эмоционально положительной состояний или подкрепление, а также формирование эмоционально отрицательной состояний очень важна для выживания и развития организма.
Наше осознанное “Я” оказывается в центре нашего субъективного мира, в середине окружающей среды, в которой мы существуем. При этом очень интенсивно работает наша память, которая превращает окружающий нас мир в наш внутренний мир, со всеми вытекающими из этого последствиями. В этот процесс очень большой вклад вносят зеркальные нейроны, которые тоже локализованы в коре.
Таким образом, зеркальные нейроны – это нейроны подражания, они активируются как при наблюдении за поведением, так и при выполнении поведения. Зеркальным нейронам приписываются следующие функции: понимание и предсказание намерений, мотивация и эмоция подражания, облегчение обучения, обучение через подражание или обучение с учителем, эмпатия, сочувствие, сопереживание, автоматическая имитация, моторная имитация, мимика, аутизм, самосознание человека, теория разума или понимание чужого сознания, речь. Все перечисленные функции относятся к субъективным ощущениям.
Вес уверенности: Имеет дополнительные подтверждения независимых специалистов
Зеркальные нейроны – это нейроны подражания, они активируются как при наблюдении за поведением, так и при выполнении поведения. Зеркальным нейронам приписываются функции, которые относятся к субъективным ощущениям, в том числе и процессы самосознание человека. Переход объективных процессов в субъективные ощущения не описывается физиологически, переход описывается через виртуальный скачок в психологию. Зеркальных клеток очень мало, они разбросаны по коре мозга маленькими островками, но их функции вовлекают большие нейронные сети. Проблему можно решить, если предположить, что зеркальные нейроны функционируют как нейроны “бабушки”. В этом случае функции, которые приписываются зеркальных нейронов, превращаются в функции коры больших полушарий, которая является частью эмоционально насыщенной когнитивной карты мозга. Критиков теории зеркальных нейронов довольно много, они считают, что нет прямых доказательств относительно предполагаемых функций зеркальных нейронов. Они предполагают, что в описанных процессах присутствует “Я” наблюдателя, что прямо декларируется в виде самосознания человека. В обзоре описывается два процесса само-опознания и само-идентификации. Процессы осознания и идентификации – это взаимодействия “Down Up” сигнала с “Top Down” сигналом из префронтальной коры. Эмоционально насыщенная когнитивная карта мозга формируется интеграцией нейронов гиппокампа нейронами коры больших полушарий. Наше “Я”, через процессы само-осознание, само-идентификацию и виртуальный скачок взаимодействуют с нейронами больших полушарий, получающих информацию о процессах в окружающей среде и формирующих контекст окружающей среды, представляющий собой наше субъективное ощущение. Новая информация о процессах в окружающей среде, консолидируется эксплицитной памятью с помощью специфической модификации экспрессии генов, и хранится в открытом доступе, то есть в состоянии быстрого воспроизведения при напоминании.
ВВЕДЕНИЕ
В течение последних нескольких лет наша лаборатория занималась проблемой “Объединенного функционирования мозга”. У нас сложилось представление о том, что модификация любого параметра, любого компонента функционирующего мозга, модифицирует весь мозг и его вновь выработанная специфическая активность консолидируется. Правда, нужно оговорится, что экспериментально это не всегда приводит к достоверно значимым результатам, особенно когда исследуются такие тонкие процессы как обучение и память. Но концепция “Объединенного функционирования мозга” очень хорошо проявляется при развитие психопатологических состояний, зависимых от возраста, которые со временем усиливаются, расширяются и втягивают другие системы и состояния. Другое название данной проблемы, это “Введение в молекулярную психонейрофизиологию”. Нетрудно понять, что проблема очень большая, но приведенный здесь обзор описывает один из центральных механизмов концепции.
Все процессы мозга можно рассмотреть с позиции реализации поведения, с позиции реализации целенаправленного эмоционально мотивированного поведения на основе соматотопии. Основными звеньями регуляции моторного поведения являются сенсомоторная кора, базальные ганглиии, красное ядро, мозжечок. Каждая из этих структур имеет свою специфику в анализе периферической афферентации и формировании моторной команды, которая реализуется через кортикоспинальную нисходящую систему мозга. Описана сенсомоторная соматотопия [7] и соматотопия экстрапирамидной системы мозга, базальных ганглиев [89].
Поведение реализуется иерархическими системами мозга [3, 4, 7] (рис. 1). Соматотопия представляет собой строго специфическую двустороннюю анатомическую связь между мышцами и различными участками моторной и соматосенсорной коры мозга. Соматотопия, это представительство всего тела в коре мозга [84].
Иерархические системы мозга функционируют следующим образом. Активность из коры приходит к определенным частям хвостатого ядра. Оттуда к определенным частям внутреннего и внешнего сегментов globus pallidus (бледный шар) и ретикулярной части черной субстанции (рис. 1). В черной субстанции информация расщепляется на три части; одна часть запускает таламокортикальные сети и возвращает информацию в кору и дозальный стриатум. Это реверберирующие, циклически функционирующие таламокортикальные сети. Вторая часть, формирует выход из базальных ганглий и вместе с красным ядром дополняет информацию, идущую из моторной коры. Третья часть, через глобальный дофаминергический (ДА) сигнал, запускает мезолимбическую, мезокортикальную и ниграстриатную ДА систему, которая через взаимодействие и интеграцию с норадренергической (НА) и серотонинергической (СТ) системами, запускает мезолимбические и мезо-кортикалные структуры и индуцирует эмоционально мотивационные состояния. По современным представлениям [2, 3] эмоционально мотивационные состояния формируются мезо-лимбическими структурами мозга: ДАергическая вентральная тегментальная область (ВТО), гиппокамп, миндалина, хабенула, таламус и прилежащее ядро; и контролируется мезо-кортикальной системой – префонтальная кора. Ключевыми системами функционирующими в этом круге, являются моноаминергические (ДА, НА, СТ) системы мозга.

Моноаминергические системы мозга, это модуляторные системы, не генерирующие ВПСП или ТПСП, так как ее рецепторы не ионотропные, а метаботропные, связанные с G белками и не несут в своем составе ионный канал. Синаптические и несинаптические системы принципиально отличаются друг от друга [16, 90]. Медиаторные ГАМК и глутаматные синаптические системы реализуют нейросетевые процессы, несинаптические нейромодуляторные системы, в частности моноаминергические системы, формируют эмоциональные и мотивационные состояния.
Пищевые, питьевые и половые центры локализованы в гипоталамусе [2]. Лептин и грелин, гормоны сытости и голода, контролируют пищевую потребность. Гормон вазопрессин контролирует концентрацию воды, осмолярность и потребность пить. Половые гормоны контролируют сексуальную потребность. Рецепторы всех этих гормонов экспрессируются в ДА нейронах ВТО, терминали которых проходят через гипоталамус.
Взаимодействие рецепторов этих гормонов с ДА системой вызывает мотивацию, которая реализуется как ГАМК сигнал из прилежащего ядра к вентральным сегментам паллидума (рис. 1). Дорзальные сегменты паллидума получают ГАМК сигналы из дорзального стриатума и выполняют целенаправленное эмоционально мотивирование поведение [2–4], удовлетворяя или не удовлетворяя потребность и мотивацию. Удовлетворение мотивации вызывает эмоционально положительное состояние и соответствующую реакцию. Неудовлетворенная мотивация вызывает эмоционально отрицательное состояние и соответствующую реакцию. Эмоционально положительное состояние, это состояние, которое направлено на собственное продление и усиление, на продление и усиление сигналов, индуцирующих эмоционально положительное состояние. Эмоционально отрицательное состояние, это состояние, которое направлено на собственное ослабление или прекращение, или на ослабление и прекращение сигналов, индуцирующих эмоционально отрицательное состояние. [4]. Подкрепление это не кусочек сахара, которое получает животное, а эмоционально положительное состояние индуцированное этим кусочком.
Но целенаправленное поведение не может быть реализовано без ориентации в окружающей среде, которая обеспечивается когнитивной картой мозга.
Когнитивную карту мозга формируют нейроны гиппокампа: эти нейроны интегрируются со зрительными, слуховыми, обонятельными и другими системами мозга и формируют контекст окружающей среды [4] (подробнее см. ниже).
Новое поведение реализуется вновь сформированной нейронной сетью. Эту нейронную сеть можно назвать энграммой памяти, и она реализуется на основании процессов синаптической пластичности. Процесс синаптической пластичности можно записать в виде: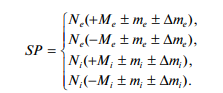
Здесь SP – эффективность синаптической пластичности, N – нативный рецептор, e – глутаматные рецепторы (excitation, возбуждение), i – ГАМКА рецептор (inhibition, торможение), M – кластеризация рецепторов, –М – декластеризация рецепторов, m – модификация субъединичной композиции рецепторов, Δm – сплайс варианты или редактирование рецепторов [6, 7].
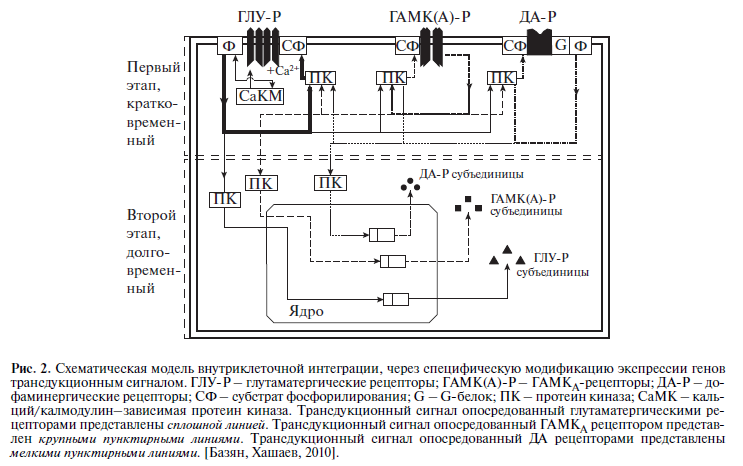
Таким образом, целенаправленное поведение реализуется с помощью нейронной сети, которая контролируется и управляется модуляторными системами мозга, формирующими состояния. Управление сетью осуществляется за счет внутриклеточной интеграции медиаторных и модуляторных систем (рис. 2).
ЗЕРКАЛЬНЫЕ НЕЙРОНЫ, ФИЗИОЛОГИЧЕСКАЯ РОЛЬ И ОСОБЕННОСТИ ФУНКЦИОНИРОВАНИЯ
Предполагается, что зеркальные нейроны имеют принципиальное значение для функционирования мозг, так как им приписывается ключевая роль в реализации высших интегративных функциях мозга. Зеркальные нейроны были открыты у обезьян в лаборатории Giacomo Rizzolatti (Джакомо Ризолетти), в университете Пармы (Италия), в середине 90-тых годов [36, 48] и изучены сотрудниками этой лаборатории . Эти нейроны отличаются тем, что они активируются как при выполнении определенного поведения, так и при наблюдении за поведением. Из-за этого они были названы зеркальными нейронами, отражающими то, за чем наблюдаешь. Зеркальные нейроны были обнаружены у макак, в нижней лобной извилине (зоне F5), затем в нижней теменной доле (поле 40) [52, 128]. Этих нейронов относительно мало 10–17% от общего числа нейронов зоны F5 [52, 63, 128].
Несколько позднее зеркальные нейроны были обнаружены у людей [15, 24, 43, 66, 67, 100, 109, 117, 134]. Когда смотришь на карту мозга, где определены зеркальные нейроны у обезьян и людей [100], то видно, что они разбросаны по всей коре больших полушарий маленькими островками (см. табл. 1). Из табл. 1 видно, что зеркальные нейроны у человека представлены более широко чем у обезьян [100].
Открытие зеркальных нейронов возродило почти забытое представление о том, что движение связано с пониманием [63]. Моторные теории когнитивности оставили длительный след в психологии [85], восходящий по крайней мере к началу XVIII в., к моторной интерпретации глубины восприятия [21], и были предложены в качестве объяснения широкого спектра психических процессов. В начале ХХ-го в. была предложена моторная теория умственных образов [137, 138], а Уотсон [139] считал, что мысль можно объяснить речевыми сенсорно-моторными процессами. Роль зеркальных нейронов [35, 39, 47, 60, 63, 108–111] анализируется как одна из последних в этой длинной списке моторных теорий, вплоть до моторной теории восприятия речи [75].
ВОЗМОЖНЫЕ ФУНКЦИИ ЗЕРКАЛЬНЫХ НЕЙРОНОВ
Многие исследователи связывают высшие интегративные процессы с функцией зеркальных нейронов.
ПОНИМАНИЕ, ПРЕДСКАЗАНИЕ НАМЕРЕНИЙ
Была зафиксирована активность зеркальных нейронов в нижней теменной доле двух макакрезусов, всего 41 нейрона. Нижние теменные доли уже давно признаны ассоциативной корой, которая объединяет сенсорную информацию. Обезьяны наблюдали, как экспериментатор либо хватает яблоко и подносит его ко рту, либо хватает предмет и ставит его в чашку В общей сложности 15 зеркальных нейронов энергично сработали, когда обезьяна наблюдала движение “схватить, чтобы съесть”, но не обнаруживали никакой активности, пока экспериментатор хватал яблоко и просто держал в руке. Других 4 зеркальных нейрона активировались в ответ на то, что экспериментатор в конце концов помещал яблоко в чашку, но не съел его. Только тип действия, а не кинематическая сила, с которой моделируются объекты, определяли активность нейронов. Также было важно, что нейроны срабатывали до того, как обезьяна видела поведение человека, запускающего второй моторный акт – поднесение объекта ко рту или помещение его в чашку. Следовательно, нейроны нижней теменной доли кодируют одно и то же действие (хватание) по-разному в соответствии с конечной целью действия [42].
Они могут предоставить нейронную основу для прогнозирования последующих действий другого человека и определения намерения.
ОБЛЕГЧЕНИЕ ОБУЧЕНИЯ
Другая возможная функция зеркальных нейронов – это облегчение обучения. Зеркальные нейроны кодируют (не реально, а виртуально) конкретное представление действия, то есть, представление, которое активировалось бы, если бы наблюдатель действовал. Это позволило бы нам моделировать (внутренне повторять) наблюдаемое действие неявно (виртуально в мозге), чтобы быстрее активировать наши собственные двигательные программы наблюдаемых действий и подготовиться к воспроизведению действия, через некоторое время. Это неявное обучение. Благодаря этому, наблюдатель будет реализовать свое действие с большей ловкостью [70].

ОБУЧЕНИЕ ЧЕРЕЗ ПОДРАЖАНИЕ ИЛИ ОБУЧЕНИЕ С УЧИТЕЛЕМ
Обучение с помощью зеркальных нейронов лучше назвать “Обучение через подражание или обучение с учителем”. Для подтверждения этого термина можно привести огромное количество примеров, не только на людях, но и на животных.
Когда родители обучают детенышей охотиться, то детеныши наблюдают как это делают родители и подражая им сами обучаются охотиться.
ЭМПАТИЯ Несколько исследователей [32, 33, 49–51, 103], независимо друг от друга утверждали, что система зеркальных нейронов вовлечена в эмпатию, сочувствие, или сопереживание. Большое количество экспериментов с использованием МРТ, ЭЭГ и МЭГ показали, что определенные области мозга (в частности, передняя инсула, передняя поясная извилина коры головного мозга и нижняя лобная кора) активны, когда люди испытывают эмоции (отвращение, счастье, боль и т.д.) и когда они видят другого человека, испытывающего эмоции [22, 74, 79, 87, 140]. Была выдвинута идея, что зеркальные нейроны имеет решающее значение для эстетического восприятия [44].
МОТИВАЦИЯ И ЭМОЦИЯ ПОДРАЖАНИЯ
В интернете я наткнулся на рассказ, в котором описывалась ситуация, как были открыты зеркальные нейроны. Сейчас этот рассказ я не могу найти, и я думаю, что это выдуманная история, но она очень хорошо описывает функции зеркальных нейронов. В лаборатории Джакомо Риззолетти исследовали поведение обезьяны, ввели электроды в нейроны ответственные за сгибание локтя и артикуляцию губ. Ввели, зафиксировали голову обезьяны и электроды. Электрические ответы нейронов вывели на монитор компьютера. При срабатывании нейрона монитор показывал электрический потенциал нейрона, но еще и щелкал. Все поставили, зафиксировали, закрепили и ушли обедать. Первым вернулся молодой сотрудник с мороженным в руках. Подошел к установке и начал рассматривать, все ли на месте, не отошел ли какой-нибудь контакт? А обезьяна следила за ним глазами. Парма, лето, жара и в некий момент молодой человек лизнул мороженное. Треск и активность мониторов ввергла его в шок. Это сработали зеркальные нейроны, которые отражали мотивацию обезьяны “я тоже хочу, и мне дай”.
Выше я описывал, что эмоциональные и модуляторные состояния формируются модуляторными, не синаптическими моноаминергическими системами мозга (НА, ДА, СТ) при высвобождении из не миенилизированных терминалей ядер среднего мозга (черная субстанция, ядро Раффе и голубое пятно). Зеркальные нейроны локализованы в коре и активируют синаптические ГАМК и ГЛУ системы коры мозга. Как было описано выше (см. рис. 1), активность из коры мозга, через стриатум и сегменты бледного шара, по нисходящей, активируют черную субстанцию, ядра среднего мозга и усиливают выделение моноаминов, которые в свою очередь, формируют эмоциональные и мотивационные состояния [2]. Получается, что источник эмоции подражания в коре, но этот источник запускает эмоциональные и мотивационные состояния через ядра моноаминергических системы среднего мозга.
Мотивация “я тоже хочу” широко распространена у людей. Мы живем в социальной среде и наблюдаем за другими людьми. Эти наблюдения вызывают у нас мотивацию подражания, “я тоже хочу”. Я тоже хочу быть успешным, я тоже хочу быть богатым, я тоже хочу иметь вот это, то и так далее. Выше я уже писал, что “удовлетворение мотивации вызывает эмоционально положительное состояние и соответствующую реакцию. Неудовлетворенная мотивация вызывает эмоционально отрицательное состояние и соответствующую реакцию”. Поэтому, когда наша мотивация “я тоже хочу” удовлетворяется, у нас проявляется эмоционально положительная реакция, к которой мы скоро привыкаем и уже не замечаем того, чего мы хотели. А когда наша мотивация не удовлетворяется, у нас возникает эмоционально отрицательная реакция. Эта не удовлетворенная мотивация может вызвать у нас чувство зависти, а чувство зависти – это страшное чувство.
АВТОМАТИЧЕСКАЯ ИМИТАЦИЯ
Термин обычно используется для обозначения случаев, когда человек, наблюдая движение тела, непреднамеренно выполняет подобное движение тела или изменяет способ, которым выполняется движение тела. Автоматическая имитация редко включает в себя открытое выполнение соответствующих ответов. Вместо этого эффекты обычно состоят из времени реакции, а не точности, различий между совместимыми и несовместимыми испытаниями. Исследования показывают, что существование автоматической имитации, которая является скрытой формой имитации, отличается от пространственной совместимости. Это также указывает на то, что, хотя автоматическая имитация подвержена входной модуляции с помощью процессов внимания и выходной модуляции с помощью тормозных процессов, она опосредована изученными долгосрочными сенсомоторными ассоциациями, которые не могут быть изменены непосредственно преднамеренными процессами [77]. Кроме того, есть данные, которые демонстрируют, что наш постуральный контроль нарушается, когда люди слушают предложения о других действиях. Например, если задача состоит в том, чтобы поддерживать осанку, люди делают это хуже, когда слушают такие предложения: “Я встаю, надеваю тапочки и иду в ванную”. Это явление может быть связано с тем, что во время восприятия действия происходит аналогичная активация моторной коры, как если бы человек совершал такое же действие (система зеркальных нейронов) [69].
МОТОРНАЯ ИМИТАЦИЯ, МИМИКА
В отличие от автоматической имитации, моторная имитация наблюдается в натуралистических социальных ситуациях и посредством измерений частоты действий в течение сеанса, а не измерений скорости и/или точности в ходе испытаний [26].
Объединение исследований по моторной и автоматической имитации может выявить правдоподобные признаки того, что эти явления зависят от одних и тех же психологических и нервных процессов. Предварительные данные, однако, получены из исследований, показывающих, что социальное праймирование оказывает аналогичное влияние на моторную имитацию [73, 136].
Тем не менее, сходство между автоматической имитацией, зеркальными эффектами и двигательной имитацией заставило некоторых исследователей предположить, что автоматическая имитация опосредована системой зеркальных нейронов и что она является строго контролируемым лабораторным эквивалентом моторной имитации, наблюдаемой в натуралистических социальных контекстах. Если это так, то автоматическая имитация может использоваться в качестве инструмента для исследования того, как зеркальная нейронная система способствует когнитивному функционированию и как двигательная имитация способствует социальным установкам и поведению [60, 96].
Мета-анализ имитационных исследований на людях позволяет предположить, что имеется достаточно доказательств активации зеркальной системы во время имитации, что вероятно участие зеркальных нейронов, хотя ни одно из опубликованных исследований не зафиксировало активность единичных нейронов. Тем не менее, это, вероятно, недостаточно для моторной имитации. Исследования показывают, что области лобной и теменной долей, которые выходят за пределы классической зеркальной системы, одинаково активируются во время имитации. Это говорит о том, что другие области, наряду с системой зеркал, имеют решающее значение для имитационного поведения [86].
К моторной имитации относится и мимика.
Мимика имеет большое значение при социальном взаимодействии. С помощью мимики мы понимаем настроение другого человека, мы понимаем какое влияние на собеседника имели наши слова или поведение. С помощью мимики мы можем понять, понимаем отношение другого человека к себе. Мимика играет большую роль при эмпатии, сочувствии и сопереживании.
АУТИЗМ
Процесс подражания и имитации вписывает нас в социальную среду. В основном, мы ведем себя соответственным образом, наблюдая за другими людьми. Возможно, что это явилось одной из причин предположения, что зеркальные нейроны связаны с аутизмом, так как аутичные больные не вписываются в социальную среду. Было высказано предположение, что проблемы с системой зеркальных нейронов могут лежать в основе когнитивных расстройств, особенно аутизма [31]. Однако связь между дисфункцией зеркальных нейронов и аутизмом является проблемной, и еще предстоит продемонстрировать, как зеркальные нейроны связаны со многими важными характеристиками аутизма [34].
Некоторые исследователи утверждают, что существует связь между дефицитом зеркальных нейронов и аутизмом. Записи ЭЭГ из моторных зон подавляются, когда кто-то наблюдает за движением другого человека, сигнал, который может относиться к зеркальной нейронной системе. Это подавление было меньше у детей с аутизмом [90].
Хотя эти результаты были воспроизведены несколькими группами [31, 97], другие исследования не нашли доказательств дисфункциональной системы зеркальных нейронов при аутизме [34].
В 2008 году было опубликовал исследование [91], в котором представлены противоречивые данные ЭЭГ. Оберман и Рамачандран нашли типичное подавление активности мю-волн для знакомых стимулов, но не для незнакомых стимулов, что привело их к выводу, что система зеркальных нейронов у детей с расстройствами аутистического спектра была функциональной, но менее чувствительной, чем у типичных детей [91]. На основании противоречивых данных, представленных в экспериментах по подавлению мю-волн, можно предположить, что результаты подавления мюволн не могут использоваться в качестве достоверного показателя для измерения производительности зеркальных нейронных систем.
ГЕНДЕРНЫЕ РАЗЛИЧИЯ
Ряд исследований, проведенных с использованием различных нейрофизиологических методов, включая магнитную энцефалографию [27], возбудимость спинального рефлекса [28], электроэнцефалографию [29, 142], подтвердили наличие гендерных различий в зеркальных реакциях человека. Нейронная система участников женского пола проявляют более сильный моторный резонанс, чем участников мужского пола.
В другом исследованиях, гендерные различия в механизмах зеркальных нейронов были подкреплены результатами, которые показали усиление эмпатической способности у женщин по сравнению с мужчинами. Во время эмоционального социального взаимодействия женщины демонстрируют большую способность воспринимать эмоциональную проекцию, чем, когда общаются с другой персоной лицом к лицу. Эта способность может быть связана с тем, что социализация мужчин в большинстве культур требует, чтобы мужчины ограничивали эмоциональную экспрессию.
Тем не менее, результаты показали, что когда дело дошло до распознавания эмоций других персон, способности мужчин и женщин были очень похожи, и не было никаких ключевых различий между ними [119].
САМОСОЗНАНИЕ ЧЕЛОВЕКА
Было сделано предположение, что зеркальные нейроны могут обеспечить неврологическую основу самосознания человека [91, 92]. В эссе, написанном для [105], Рамачандран дал следующее объяснение своей теории: “… Я также предположил, что эти нейроны могут не только помочь имитировать поведение других людей, но могут быть обращены “внутрь”– как это было для создания представлений второго порядка или мета представлений ваших собственных более ранних мозговых процессов. Это может быть нервной основой самоанализа и взаимности самосознания и другого осознания. Очевидно, что здесь возникает вопрос “курица или яйцо” относительно того, что возникло первым, но главное, что эти два представления эволюционировали, взаимно обогащая друг друга, чтобы создать зрелое представление о себе, которое характеризует современных людей” [105].
Если иметь ввиду функцию зеркальных нейронов, то в этой функции выявляется личность наблюдателя – я подражаю. Автор этой статьи, (А.С.Б) не очень понимает, что имеет ввиду Рамачандран. По моему мнению, есть два других, довольно четких описания процессов само-осознания и само-идентификации. Само-осознание или само-опознание – это зеркальное тестирование, тест метки или mirror self-recognition test (MSR), предложенный в 1970 году психологом Гордоном Галлапом Младшим [17]. Суть работы сводится к тому, что обезьянке на лбу рисуют пятно белого цвета и подводят ее к зеркалу. Увидев образ в зеркале, обезьянка тычет пальцем в пятно у себя на лбу, а не в зеркале. Этот тест на само-опознание проходят: приматы (шимпанзе, гориллы, орангутанги) и люди. Люди осознают себя заглядывая в зеркало начиная с двух лет [см. фильм 11]. Этот тест проходя так же азиатские слоны, дельфины, касатки и евразийская сорока.
Другой метод – это нейровизуализация идентичности, идентификация [10]. Человека сажают в FMRI (functional magnetic resonance imaging – изображение получаемое с помощью функционального магнитного резонанса), дают в руки пачку фотографий, в том числе и его собственную фотографию и предлагают рассмотреть их. Дойдя до своей фотографии человек идентифицирует себя. При этом резко активируется медиальная префронтальная кора. В меньшей степени в процессе участвуют, передняя и задняя поясная извилина, и область предклиньев. Это осознание себя. У двухлетнего ребенка, в этот момент возникает свое “Я” [см. фильм 11]. Процессы осознания и идентификации – это взаимодействие “Down Up” сигнала (сигнал снизу вверх – формирование образа) с “Top Down” сигналом из префронтальной коры [86, 127, 132]. Между прочим “Top Down” сигнал, это и сигнал селективного внимания. Проблема “Top Down” сигнала очень большая и она требует отдельной статьи. Наше “Я” формируется в коре мозга, а зеркальные нейроны раскиданы по всей коре мозга маленькими островками (см. табл. 1). Я полагаю, что при процессе самоидентификации активируется эмоционально насыщенная когнитивная карта мозга. Подробнее об этом я напишу ниже.
СОМНЕНИЯ ОТНОСИТЕЛЬНО ЗЕРКАЛЬНЫХ НЕЙРОНОВ
Несмотря на то, что многие ученные начали поддерживать идею зеркальных нейронов и разрабатывать теорию их функционирования, есть ученые, которые выразили сомнения как в существовании, так и в роли зеркальных нейронов у человека [63, 96]. По мнению Хикокa [63] зеркальных нейронов очень мало, например всего 10–17% от всех нейронов зоны F 5, ростральной части вентральной премоторной области. Причем эти результаты цитируются из работ авторов, которые открыли зеркальные нейроны [36, 48].
При этом статья Хикока [63] называется “Восемь проблем теории в понимании действий зеркальных нейронов у обезьян и человека”. Выступая против утверждения, что зеркальные нейроны участвуют в понимании действий, он пришел к выводу, что “ранняя гипотеза о том, что эти клетки лежат в основе понимания действий, является интересной и разумной идеей. Однако, несмотря на его широкое признание, предположение никогда не было адекватно проверено на обезьянах и людях.
Другие авторы утверждали, что первоначальные анализы были неубедительными, потому что они основывались на качественных описаниях индивидуальных свойств клеток и не учитывали небольшое количество сильно зеркально-селективных нейронов в моторных областях [35]. Другие ученые утверждают, что измерения задержки пачечной активности нейронов, по-видимому, не совместимы со стандартным временем реакции [96] и указали, что никто не сообщал, что прерывание моторных областей в F5 приведет к снижению распознавания действия [61]. (Критики (этого аргумента) ответили, что авторы пропустили нейропсихологические исследования человека, и исследования c транскраниальной магнитной стимуляцией, сообщающие, что нарушения этих областей, действительно вызывают дефицит действия [102, 134] не затрагивая другие виды восприятия [116].
В 2009 г. Лангнау с коллегами провели эксперимент, в котором они сравнили двигательные акты, которые сначала наблюдались, а затем выполнялись, с моторными актами, которые сначала выполнялись, а затем наблюдались. Они пришли к выводу, что существует значительная асимметрия между двумя процессами, которая указывает на то, что зеркальные нейроны не существуют у людей.
Они заявили: “Важно отметить, что мы не обнаружили никаких признаков адаптации для двигательных актов, которые сначала выполнялись, а затем наблюдались. Неспособность найти кроссмодальную адаптацию для выполненных и наблюдаемых двигательных актов не совместима с основным предположением теории зеркальных нейронов, которая утверждает, что распознавание и понимание действий основаны на моторной симуляции” [77]. Однако в том же году было показано, что если целевые действия используются в качестве стимулов, то, как нижняя теменная доля, так и премоторные области показывают подавление повторения между наблюдением и выполнением, которое предсказывается зеркальными нейронами [68].
Владимир Косоногов видит другое противоречие. Сторонники теории зеркальных нейронов постулируют, что зеркальные нейроны кодируют целенаправленные действия, потому что они активируются, если наблюдаемое действие является целенаправленным. Однако зеркальные нейроны активируются только тогда, когда наблюдаемое действие направлено на цель (объектно-ориентированное действие или коммуникативный жест, который, безусловно, также имеет цель). Как они “узнают”, что определенное действие направлено на достижение цели? На каком этапе их активации они обнаруживают цель движения или его отсутствие? По его мнению, зеркальная нейронная система может быть активирована только после того, как цель наблюдаемого действия прописывается некоторыми другими структурами мозга [71].
ТЕОРИЯ РАЗУМА ИЛИ ПОНИМАНИЕ ЧУЖОГО СОЗНАНИЯ
Под термином “понимание чужого сознания” (theory of mind) подразумевают способность делать выводы о всем наборе психических состояний (надежды, желания, намерения, воображения, эмоции и др.), которые являются причиной действий другого индивидуума. Понимание чужого сознания – это способность отражать сознание других индивидуумов [21]. Способность понимать намерения, связанные с действиями других индивидуумов, является фундаментальным компонентом социального поведения [47]. В настоящее время термин “theory of mind” широко используется в англоязычной литературе при объяснении механизмов обучения, актерских способностей, развития мышления и способностей к социальному взаимодействию, описании симптоматики аутизма как следствия неправильной работы зеркальных нейронов [70, 71].
“Теория ума” относится к нашей способности выводить психическое состояние другого человека (то есть убеждения и желания) из опыта или его поведения. Есть несколько конкурирующих моделей, которые пытаются объяснить теорию нашего мышления; наиболее заметным по отношению к зеркальным нейронам является теория симуляции. Согласно теории симуляции, теория разума доступна, потому что мы подсознательно сопереживаем человеку, за которым наблюдаем, и, учитывая соответствующие различия, представляем, чего бы мы делали на его месте [55, 56].
Зеркальные нейроны были интерпретированы как механизм, с помощью которого мы моделируем других, чтобы лучше понять их, и поэтому их открытие было воспринято некоторыми как подтверждение теории модирования (которая появилась за десятилетие до открытия зеркальные нейроны) [50]. Совсем недавно, Теория Разума и Симуляции была замечена как взаимодополняющие системы с различными временными курсами развития [66, 85, 125].
РЕЧЬ
У людей функциональные МРТ исследования выявили области, гомологичные зеркальной нейронной системе обезьяны в нижней лобной коре, вблизи области Брока, одной из предполагаемых языковых областей мозга [18]. Это привело к предположениям о том, что человеческий язык развился из системы представления/понимания жестов, реализованной в зеркальных нейронах. Эта гипотеза подтверждается некоторыми цитоархитектоническими гомологиями между премоторной областью F5 обезьяны и областью Брока человека [101]. Показатели расширения словарного запаса связаны с возможностями детей вокально имитировать не слова, а похожие звуки и, таким образом, приобретать новые произношения слов. Такое повторение речи происходит автоматически, быстро [103] и облегчает восприятие речи [81, 82]. Кроме того, такая вокальная имитация может происходить без понимания [41, 79].
Еще одним подтверждением этой связи является исследование, в котором активность мозга двух участников измерялась с помощью фМРТ, в то время как они жестикулировали словами друг другу с помощью жестов рук с игрой в шарады – способ, который, по мнению некоторых, может представлять эволюционный предшественник.
человеческого языка. Анализ данных с использованием причинно-следственной связи выявленной методом Грейнджера показал, что зеркальнонейронная система наблюдателя действительно отражает характер активности в двигательной системе отправителя, подтверждая идею о том, что моторная концепция, связанная со словами, действительно передается из одного мозга в другой. с использованием системы зеркал [120].
Зеркальные нейроны исследуются с помощью объективных методов: потенциалы клеток, FMRI, фМРТ, магнитная энцефалография, возбудимость спинального рефлекса, ЭЭГ и другие. В итоге в этих исследованиях получаются конкретные результаты на клеточном или структурном уровне.
Но процессы, которые описываются как функции зеркальных нейронов, в цитированной выше литературе, относятся к субъективным ощущениям.
Но переход объективных процессов в субъективные процессы не описывается, не только в цитированных выше работах, а в принципе не описываются.
ОБЪЕКТИВНЫЕ И СУБЪЕКТИВНЫЕ ПРОЦЕССЫ МОЗГА, УЧАСТИЕ ЗЕРКАЛЬНЫХ НЕЙРОНОВ
Переход объективных процессов мозга в субъективные процессы очень сложная многоуровневая задача, которая имеет виртуальное решение.
В окружающем нас физическом мире нет ни света, ни цвета (электромагнитное излучение), ни звука (вибрационные колебания в звуковом диапазоне), ни вкуса (молекулы, вкусовые рецепторы), ни запаха (летучие молекулы, обонятельные рецепторы), ни боли (физические воздействия, тактильные рецепторы), но мы их ощущаем. Более того, мы понимаем смысл услышанных слов.
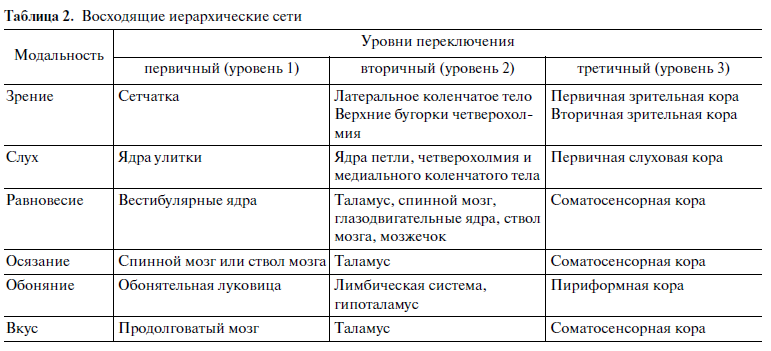
В мозге есть объективные центры этих субъективных ощущений (см. табл. 2), но переход объективных процессов мозга в субъективные ощущения не описывается [1, 11–13]. В то же время для нас субъективные ощущения самые реальные.
Для описания перехода объективных процессов в субъективные ощущения необходимо использовать “черный ящик” [1, 12]. “Черный ящик”, – это кибернетическое устройство, где известно что на входе и что на выходе, а что творится внутри – неизвестно, или неинтересно. Наш “черный ящик” осуществляет виртуальный скачок. Активируются зеркальные нейроны, потом виртуальный скачек, появляется то чего не было до сих пор: понимание чужого сознания; сочувствие; эмоции; понимание и предсказание намерений; самосознание человека; мышление, и так далее и тому подобное. Наш “черный ящик” двусторонний [1], все что творится в нашем субъективном мире, влияет на наши объективные процессы, меняет активность зеркальных нейронов, модифицирует все процессы и сохраняет (консолидирует) их.
Но есть еще одна проблема. Зеркальных нейронов очень мало. Например, выше было описано [43], что в нижней теменной доле двух макак – резусов, была зафиксирована активность всего 41 зеркального нейрона. Обезьяны наблюдали, как экспериментатор либо хватает яблоко и подносит его ко рту, либо хватает предмет и ставит его в чашку В общей сложности 15 зеркальных нейронов энергично сработали, когда обезьяна наблюдала движение “схватить, чтобы съесть”, но не обнаруживали никакой активности, пока экспериментатор хватал яблоко и просто держал в руке.
Других 4 зеркальных нейрона активировались в ответ на то, что экспериментатор в конце концов помещал яблоко в чашку, но не съел его. Сосчитано, что в мозге около 68 миллиардов нейронов [19, 59, 60, 133]. А в коре мозга мужчин около 23 миллиарда нейронов, а женщин – 19 миллиардов [59, 60, 133]. Сразу понятно, что количество зеркальных нейронов очень мало для запуска процессов в мозге. Но проблема очень легко решается если допустить, что зеркальные нейроны функционируют как нейроны “бабушки”.
ЗЕРКАЛЬНЫЕ НЕЙРОНЫ – НЕЙРОНЫ “БАБУШКИ”
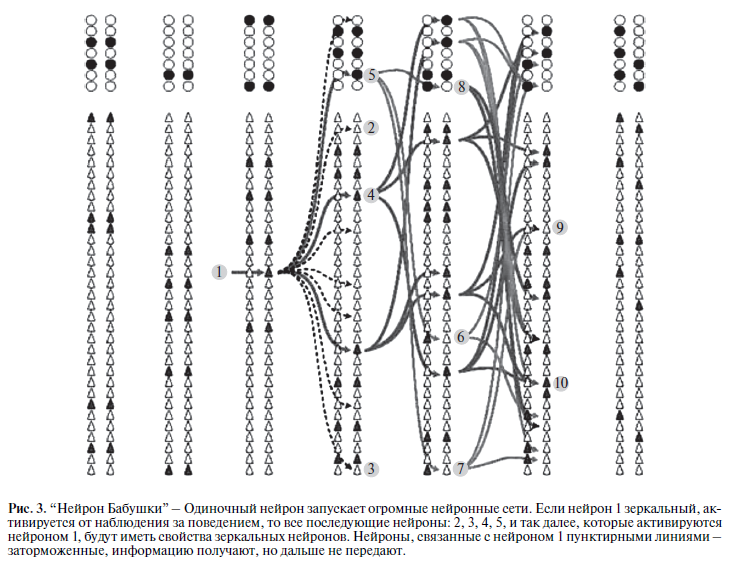
Под нейроном “бабушки” в медиальной височной доле подразумевают одиночный нейрон, активация которого вызывает активацию большого количества нейронов, точнее, сеть зрительных образов, и, следовательно, можно ожидать, что каждая клетка отвечает на многие изображения (образы) с аналогичными основными функциями [105]. Например, описывается, что при кодировании и распознавании образов, одиночные нейроны в медиальной височной доле различают лица от неодушевленных предметов [46]. Некоторые одиночные нейроны отвечают селективно на определенные эмоциональные выражения лица или объединенных показателей эмоционального выражения лица и пола. Такие одиночные нейроны особенно выявляются во время распознавания, а их ответы зависят от новизны стимула или наоборот, когда стимул уже знакомый. Следы предъявления стимула на лица или на предметы удерживались от несколько секунд до 10 часов после удаления раздражителя. Некоторые нейроны поддерживают запись предыдущего предъявления стимула, более точно, чем сознательное воспоминание человека.
Предполагается, что медиальная височная доля человека создает “когнитивную карту” параметров стимула, которая сравнима с картой (контекстом) окружающей среды, которая описана для нейронов гиппокампа грызунов [46].
В следующей работе этих авторов [72] описывается, что гиппокамп, миндалина и энторинальная коры головного мозга получают сходящиеся входы от височных регионов неокортекса, специализирующихся на обработке сложных визуальных стимулов и играют важную роль в представлении и распознавании зрительных образов. В записях нейронной активности от 427-ми одиночных нейронов в гиппокампе, энторинальной коре и миндалине человека, было обнаружено значительная степень специфической импульсации отдельных нейронов в основной последовательной цепочке. Из зарегистрированных нейронов, 14% выборочно ответили на визуальные раздражители разных категорий, в том числе лица, пейзажи и дома, известных людей и животных. На основании частоты импульсной активности отдельных нейронов, можно предсказать категорию стимула со средней вероятностью ошибки 0.24. В гиппокампе доля нейронов, отвечающих за пространственную ориентировку, схему или план была больше, чем в других структурах. Авторы считают, что эти данные обеспечивают непосредственную поддержку предположения о роли медиальных височных областей человека в представлении различных категорий визуальных стимулов [72].
В следующей работе этих авторов [105] было показано, что нейроны в верхних ступенях вентральной зрительного пути обезьян реагировали на сложные образы, такие как лица или объекты, и показывали некоторую степень инвариантности по отношению к метрических свойств, таких как размер стимула, положение и угол обзора. В этой работе сообщается о подгруппе нейронов в медиальной височной доле, которые избирательно активируются совершенно различными картинками лиц, памятников архитектуры или объектов, а в некоторых случаях даже буквами строк с их именами. Эти результаты предполагают инвариантный, редкий эксплицитный (явный) код, который может иметь важное значение в превращении сложных визуальных восприятий в долгосрочно сохраняющуюся память.
Можно предположить, что каждая клетка запускает или активирует многие образы с аналогичными основными функциями. Так как эти образы локализованы в когнитивной карте, то можно предположить, что одиночный нейрон в состоянии активировать всю когнитивную карту мозга (рис. 3). Тогда получается, что все фантастические функции, которые приписываются зеркальным нейрона, на самом деле являются функциями когнитивной корты мозга.
ЭМОЦИОНАЛЬНО НАСЫЩЕННАЯ КОГНИТИВНАЯ КАРТА МОЗГА
Как уже было написано выше, все реакции, которые приписываются зеркальным нейронам, относятся к субъективным реакциям. В такого типа реакциях ключевую роль играет осознание собственного “Я”, или представления “Я есть”.
Так же было сказано выше, что все субъективные реакции, в том числе и “Я есть”, не описываются объективными процессами, а формируются проходя через черный ящик, то есть виртуальный скачек. Но все объективные процессы до черного ящика можно описать, и в этом самое активное участие принимает эмоционально насыщенная когнитивная карта мозга.
КОГНИТИВНАЯ КАРТА МОЗГА
Ранее мы описывали эмоционально насыщенную когнитивную карту мозга с позиции целенаправленного эмоционально мотивированного поведения [4, 5], и c позиции управления синаптической пластичностью [6, 7]. Сейчас, мы более подробно опишем строение эмоционально насыщенной когнитивной карты мозга, ее функций с позиции формирования представления “Я есть”, и с позиции функций зеркальных нейронов. Идею когнитивной карты мозга выдвинул Толман в 1948 г.
[131]. Эту идею подтвердил О'Кииф, открытием “клеток места” [94] и связав ее с когнитивной картой мозга [95]. За открытие в мозге млекопитающих “клеток места” и клеток, отвечающих за ориентацию в пространстве – “навигационные клетки” – в 2014 г. Джон О'Киф, Эдвард Мозер и Мэй-Бритт Мозер получил Нобелевскую премию по физиологии и медицине. Клетки места гиппокамп – это специальные клетки в каждом из слоев структуры, которые разряжаются комплексной пачечной активностью, когда животное движется через определенное место в окружающей среде (рис. 4а). Область, в которой клетки разряжаются с наибольшей частотой, это “частотное поле” или “поле места”. Внутри этого поля нейроны места могут показать максимальную частоту разрядов 20 Гц или более, вне этого поля эти нейроны разряжаются частотой 0.1 Гц. В связи с достаточным количеством клеток места, их поля в состоянии картировать или покрывать любую данную окружающую среду. Предполагается, что эти нейроны используются не только для ориентации в пространстве, а также формируют когнитивную карту, так как в дальнейшем эти нейроны вовлекаются в процессы памяти и на них могут накладываться существенные элементы и эпизоды опыта.
Если, например, накормить животное в оном из рукавов восьмирукавного лабиринта, то у животного в 200 раз усилится активность нейрона гиппокампа ответственного за это место. Когда животное выходит их этого рукава, активность этого нейрона возвращается на контрольный уровень.
Если через некоторое время животное возвращается, входит в этот рукав лабиринта и подходит к месту кормления, то активность нейрона опять усиливается, несмотря на то, что еды там нет [129]. Эта срабатывает память на то, что здесь крысу кормили.
Большую роль в формировании когнитивной карты играют навигационные нейроны, или “нейроны сетки” (рис. 4б), локализованные в энторинальной коре и открытые супругами Мозер и их учениками в 2005 году [57]. Энторинальная кора – основной вход для нейронов гиппокампа.
Нейроны сетки разряжаются в ответ на свободное движение животного, и перекрывают пространства, которые примерно равны по размеру и расположены в периодическом треугольнике, покрывающем все доступное поле, в котором движется животное. То есть, навигационные нейроны своими разрядами “обстреливают” контролируют, точнее регистрируют не сами “нейроны места”, а “частотное поле” или “поле места”. Периодический паттерн разрядов возникает независимо от конфигурации ориентиров, как в темноте, так и в присутствии видимых ориентиров, независимо от скорости и направления движения [57]. Тут необходимо отметить, что крысы – ночные животные, у них слабое зрение, они лучше ориентируются по запаху.
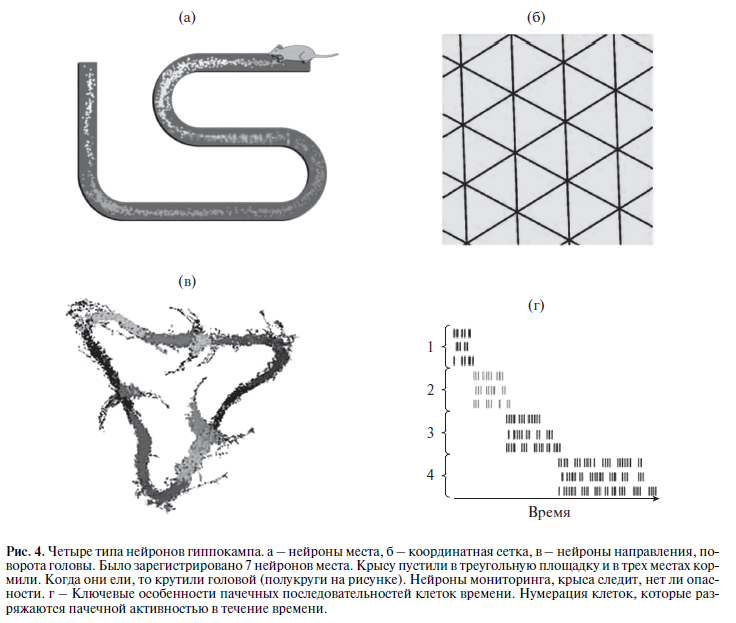
Также в энторинальной коре обнаружены нейроны направления, которые зависят от поворота головы [115]. Они играют принципиальную роль при формировании когнитивной карты. В экспериментальных условиях эти нейроны активируются, когда голова животного отворачивается от исходного направления на 45°. При таком повороте, вероятно, образ восприятия окружающего пространства меняется: и обонятельного пространства, и зрительного, и слухового и так далее.
Но энторинальная кора не единственное место локализации этих нейронов. Они обнаружены также в других областях головного мозга, в том числе: пост-субикулюм, ретроспленальная кора, таламус (передние и боковые дорзальные ядра таламуса), боковое маммилярное ядро, дорзальное тегментальное ядро, стриатум [115]. Фактически нейроны поворота головы осуществляют “мониторинг” окружающей среды и вместе с нейронами места и навигационными нейронами формируют целостный образ окружающей среды (рис. 4в) Исследования выявили существование нейронов времени в гиппокампе, которые реагируют активацией в ответ на протекающие друг за другом последовательные моменты в течение опыта [38].
Автор считает, что такое временное кодирование не может быть соотнесено с внешними событиями или пространственными измерениями опыта. Как полагает автор, эти нейроны времени представляют собой течение времени в конкретных воспоминаниях, и поэтому получил название “клетки времени”. Импульсные свойства клеток времени совпадают с импульсными свойствами “клеток места” гиппокампа. Таким образом, “клетки времени” обеспечивают дополнительное измерение, интегрированное с пространственным отображением.

Прочное представление времени и пространства в гиппокампе предполагает фундаментальный механизм для формирования пространственно-временного континииума в рамках согласованных воспоминаний. Можно полагать, что клетки времени, совпадающие по особенностям функционирования с клетками места, выстраивают внешние события во временной последовательности, так как активность клеток места меняется и при изменении внешних событий. Это очень важно, так как память консолидирует временную последовательность событий, и память воспроизводится во временной последовательности. Кстати, работа этого автора, изданная в 2013 году, так и называется “Память на время” [37].
Современная физика определяет соотношение пространства и времени как пространственновременной континуум, то есть непрерывность, неразрывность пространства и времени. Мы также воспринимаем непрерывность нашего трехмерного пространства (высота, ширина и глубина) и его неразрывность со временем, которая ограничена длительностью нашей жизни. Это неразрывность воспринимается лично каждым из нас и реализуется с помощью нашей памяти. Мы запоминаем и воспроизводим события в определенной последовательности и связываем эти события с определенным местом действия.
Летом 2015 г. c разницей в пять дней, были напечатаны работы из двух различных лабораторий, в которых описаны нейроны скорости в медиальной энторинальной коре. Особенности этих нейронов состоят в том, что при повышении скорости бега, у крыс повышается частота разрядов этих нейронов [69, 126]. Все эти клетки гиппокампа и энторинальной коры формируют сеть с нейронами различных областей коры больших полушарий и становятся основой когнитивной карты мозга (рис. 5).
ЭМОЦИОНАЛЬНО НАСЫЩЕННАЯ КОГНИТИВНАЯ КАРТА МОЗГА
Можно полагать, что “поле места” формируется под влиянием эмоционально-мотивационного состояния, когда животное получает пищу в одном из рукавов восьми-рукавного лабиринта, или когда крыса находит платформу в лабиринте Мориса и выходит из воды.
Объединение функций или совместное рассмотрение (рис. 1, 4 и 5) организует, так называемую, эмоционально насыщенную когнитивную карту мозга. Когнитивная карта становится эмоционально насыщенной, так как гиппокамп тесно контактирует с миндалиной, одной из эмоциогенных структур мозга [39, 42, 142]. Рис. 1 показывает, что любой стимул или сигнал, даже одиночный, проходящий через экстрапирамидную систему, эмоционально оценивается [4, 5].
Хорошо это или плохо, усилить этот сигнал или ослабить. А через экстрапирамидную систему проходит вся нисходящая от коры информация. Когда информация совершенно новая и поступает впервые, то выработанная оценка, или реакция, или состояние консолидируется вместе со стимулом или сигналом, как описано на рис. 2, сохраняется в памяти и используется в дальнейшем. При взаимодействии системы, представленной на рис. 1, с системой, представленной на рис. 4 и 5, происходит эмоциональная оценка когнитивной карты, точнее, образов когнитивной карты и возникает эмоционально насыщенная когнитивная карта. Оценочная функция определяет полезно или вредно то или другое явление, процесс или образ, усилить его или избавиться от него. Нужно отметить, что кора и именно оценочная функция интенсивно используется при реализации целенаправленного поведения. Целенаправленное поведение это активный поиск объектов удовлетворения потребности, которая контролируется когнитивной картой и подкрепляется или не подкрепляется оценочной функцией эмоциональных состояний.
Оценочная функция, точнее формирование эмоционально положительной состояний или подкрепление, а также формирование эмоционально отрицательной состояний [2] очень важна для выживания и развития организма. Как видно из рис. 1, оценочная функция или эмоциональные состояния формируются мезо-лимбическими и мезо-кортикальными структурами мозга: ВТО, гиппокамп, миндалина, хабенула, гиппоталамус, прилежащее ядро и префронтальная кора. Но предполагается и другая система формирования эмоциональных и мотивационных состояний – это круг Пейпеца, в которой гиппокамп занимает ключевое положение.
ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЯ “Я ЕСТЬ” И ЭМОЦИОНАЛЬНО НАСЫЩЕННАЯ КОГНИТИВНАЯ КАРТА МОЗГА, ВЗАИМОДЕЙСТВИЕ С ЗЕРКАЛЬНЫМИ НЕЙРОНАМИ
В таблице 2 описаны восходящие иерархические сети. В конечном счете это восходящие иерархические сети мозга, так как они кончаются в коре мозга. На самом деле это сенсорные сигналы, которые имеют разную модальность, идут от наших периферических органов и заканчиваются в коре [12]. С помощью этих восходящих иерархических сетей мы видим слышим, чувствуем и так далее, и в конечном счете, познаем окружающий нас мир.
Ранее было описано, что формирование представления “Я есть” начинается в 2 года, с помощью теста зеркала. У нас в зрительной коре формируется наш образ, который взаимодействует с Top Dawn сигналом из префронтальной коры. Но кроме зрительной системы, мы получаем информацию от всех органов чувств. Они тоже формируют образы и тоже идентифицируются Top Dawn сигналом из префронтальной коры. То есть в представлении “Я есть” участвуют все органы наших чувств. Но вся кора входит в состав эмоционально насыщенной когнитивной карты мозга.
Наше осознанное “Я” оказывается в центре нашего субъективного мира, в середине окружающей среды, в которой мы существуем. При этом очень интенсивно работает наша память, которая превращает окружающий нас мир в наш внутренний мир, со всеми вытекающими из этого последствиями. В этот процесс очень большой вклад вносят зеркальные нейроны, которые тоже локализованы в коре.
Что такое эмоционально насыщенная когнитивная карта, взаимодействующая с зеркальными нейронами и включающая в себя наше “Я”? Это внутренний источник нашей психики, нашего опыта и наших знаний. Если определить когнитивность, как накопление опыта и знаний и передача накопленного опыта и знаний другим особям или субъектам, то контекст окружающей среды становится внутренним источником нашего опыта, знаний и психики.
Можно ли описать проявления эмоционально насыщенной когнитивной карты. Это очень сложно, потому что нам придется описать всю нашу жизнь и все наши чувства. В нашу эмоционально насыщенную когнитивную карту входит:
все проблемы, восторги и разочарования; все наши поездки, все наши встречи с незнакомыми людьми и друзьями; мы не перестаем учиться, и накапливаем новый опыт и новые знания. Вся эта информация консолидируется, сохраняется и воспроизводится при взаимодействии структур мозга, и молекулярно-химических процессов приведенных на этих четырех схемах (рис. 1, 2, 4, 5). Самое интересное, что для формирования эмоционально насыщенной когнитивной карты и мы и животные, в основном, используем один вид памяти: увидел–запомнил, услышал–запомнил, почувствовал запах–запомнил. Обычно этот процесс называют декларативной памятью. Но если обратится к официальному термину декларативной памяти, то она гораздо шире. В реальности декларативная память описана для людей и включает в себя вербальную компоненту, которая свойственна только людям.
Декларативная память иногда называют эксплицитной (явной) памятью, которая является одним из двух типов долгосрочной памяти. Декларативная память относится к памяти, которая может быть сознательно воспроизведена таким то фактом или знанием [136]. Декларативная память это память, о которой человек может заявить на словах, в то время как эксплицитная (явная) память, это преднамеренное воспоминание информации, которую человек признает в качестве памяти [134].
Альтернатива декларативной памяти известна как не декларативная или процедурная память, которая включает бессознательные воспоминания, такие как навыки, например, обучение езды на велосипеде, обучение плавать. Декларативную память можно разделить на две категории: эпизодическую память, которая хранит конкретные личные переживания, и семантическую память, которая хранит фактическую информацию [65].
Предполагается, что обучение при декларативной памяти не требует награды, которая некоторыми авторами приравнивается к подкреплению [108, 121, 122, 130] и из этого можно сделать вывод, что декларативная память не нуждается в подкреплении. Но как было описано в цитированных ниже работах, в подкреплении нуждаются даже оборонительные рефлексы. Подкрепляется избавления затем избегания неприятного воздействия [26, 58, 107, 114, 121, 122]. Из этого можно сделать вывод, что подкреплением является эмоционально положительное состояние, которая формируется в мезо-кортико-лимбическо ДА системе мозга [2]. Награда также вызывает эмоционально положительное состояние в мезокортико-лимбическо ДА системе и подкрепляет обучение. В этом случае любой вид обучения, в том числе и декларативное, увидел–запомнил, услышал–запомнил, так же подкрепляется, так как в любом случае вызывает эмоциональную реакцию через черную субстанцию (рис. 1). Под контестом окружающей среды я понимаю вес комплекс субъективных ощущений, который сформирован у каждого из нас.
ЗАКЛЮЧЕНИЕ
Таким образом, зеркальные нейроны – это нейроны подражания, они активируются как при наблюдении за поведением, так и при выполнении поведения. Зеркальным нейронам приписываются следующие функции: понимание и предсказание намерений, мотивация и эмоция подражания, облегчение обучения, обучение через подражание или обучение с учителем, эмпатия, сочувствие, сопереживание, автоматическая имитация, моторная имитация, мимика, аутизм, самосознание человека, теория разума или понимание чужого сознания, речь. Все перечисленные функции относятся к субъективным ощущениям, особенно самосознание человека, а сами зеркальные нейроны исследуются объективными методами. Переход объективных процессов в субъективные ощущения не описывается физиологически, переход описывается через виртуальный скачек в психологию.
Зеркальных клеток очень мало, они разбросаны по коре мозга маленькими островками, но их функции вовлекают большие нейронные сети.
Проблему можно решить, если предположить, что зеркальные нейроны функционируют как нейроны “бабушки”. В этом случае функции, которые приписываются зеркальным нейронам, превращаются в функции коры больших полушарий, которая является частью эмоционально насыщенной когнитивной карты мозга.
Критиков теории зеркальных нейронов довольно много, и вся критика относится к тому, что нет прямых доказательств относительно предполагаемых функций зеркальных нейронов.
На основании этого делается вывод: что в описанных процессах присутствует “Я” наблюдателя, что прямо декларируется в виде самосознания человека. Известно два типа экспериментов исследования самосознания. Первый – это тест зеркала или тест само-опознания, а второй – тест само-идентификации. Второй тест реализуется в FMRI и выявляет значительную активацию медиальной префронтальной коры. Процессы осознания и идентификации – это взаимодействия “Down Up” сигнала (сигнал снизу вверх – формирование образа) с “Top Down” сигналом из префронтальной коры. Сигналы “Down Up” формируются восходящими иерархическими сетями мозга, которые связывают периферические реакции зрения, слуха, тактильных реакций и так далее, через подкорковые структуры с корой больших полушарий. После формирования образа, зрительного, слухового и так далее, “Top Down” сигнал префронтальной коры накрывает этот образ, происходит идентификация или осознание образа через виртуальный скачек.
Эмоционально насыщенная когнитивная карта мозга формируется интеграцией нейронов гиппокампа с корой больших полушарий. Четыре типа клеток гиппокампальной формации: клетки места, клетки времени, клетки мониторинга и клетки сетки или навигационные нейроны интегрируются с клетками коры больших полушарий и формируют когнитивную карту мозга. Гиппокамп взаимодействует с мезо-лимбическими и мезо-кортикальными структурами и формирует эмоционально насыщенную когнитивную карту мозга. Наше “Я”, через процессы само-осознания и само-идентификации и виртуальный скачек взаимодействуют с нейронами больших полушарий, получающих информацию о процессах в окружающей среде и формируют контекст окружающей среды, представляющий собой наше субъективное ощущение. Новая информация о процессах в окружающей среде, консолидируется эксплицитной памятью с помощью специфической модификации экспрессии генов, и хранится в открытом доступе, то есть в состоянии быстрого воспроизведения при напоминании.
Работа выполнена при поддержке программ РАН и РФФИ, проект № 17-29-01005-офи_м.
СПИСОК ЛИТЕРАТУРЫ 1. Базян А.С. Воззрения Л.М. Чайлахяна на истоки происхождения сознания и психики // Биофизика. 2010. Т. 55. № 3. С. 557–562.
2. Базян А.С., Рогаль А.В. Нейрохимические механизмы возникновения потребности, мотивации и целенаправленного поведения // Успехи физиол.
наук. 2015. Т. 46. № 1. С. 3–21.
3. Базян А.С. Мотивационные и эмоциональные состояния: структурные, cистемные, нейрохимические, молекулярные и клеточные механизмы // Успехи физиол. наук. 2016а. Т. 47. № 1. С. 15–33.
4. Базян А.С. Интеграция высшие функции мозга, как основа когнитивности // Успехи физиол. наук. 2016б. Т. 47. № 3. С. 19–23.
5. Базян А.С. Молекулярные, клеточные и системные механизмы мотивационных и эмоциональных состояний, реализация целенаправленного поведения и эмоционально насыщенная когнитивная карта мозга // XVIII Международная научно-техническая конференция “Нейроинформатика – 16": Лекции по нейроинформатике. М:
НИЯУ МИФИ 2016. С. 147–170. 224 с.
6. Базян А.С. Процессы синаптической пластичности на дендритном дереве нейрона // Труды конгресса по интеллектуальным системам и информационным технологиям “IS&IT’18”. Научное издание в 3 томах. Таганрог: Изд-во Ступеина С.А.
2018. Т. 2. С. 320–327.
7. Базян А.С. Соматотопия структур мозга, реализация целенаправленного эмоционально мотивированного поведения и энграмма памяти // Усп.
физиол. наук. 2019. Т. 50. № 1. С.
8. Блюм Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение. М.: Мир, 1988. С. 248.
9. Косоногов В. Зеркальные нейроны: краткий научный обзор. Ростов-на-Дону, 2009. с. 24.
http://lit.lib.ru/k/kosonogow_w_w/kosonogow-zerkalxnyenejronykratkijnauchnyjobzordoc.shtml.
10. Мельников М.Е., Штарк М.Б. Нейровизуализационные исследования идентичности // Успехи физиол. наук. 2015. Т. 46. № 4. С. 15–27.
11. Перед зеркалом. 2018. https://www.facebook.com/kimbrough.
pace/videos/10216220515721652/.
12. Смит, К.Ю.М. Биология сенсорных систем. М.:
БИНОМ. Лаборатория знаний, 2005. 583 с.
13. Чайлахян Л.М. Истоки происхождения психики, или сознания. Пущино. 1992.
14. Чайлахян Л.М. Проблема индивидуального сознания // Успехи физиол. наук. 2009. Т. 54. № 2. С.
15. Acharya S., Shukla S. Mirror neurons: Enigma of the metaphysical modular brain // J. Nat. Sci. Biol. Med.
2012. V. 3. № 2. P. 118–124.
16. Alexander S.P.H., Mathie A., Peters J.A. Guide to Receptors and Channels (GRAC), 5th edition // Brit. J.
Pharmacol. 2011. V. 164. Sappl. 1. P. S1–S324.
17. Anderson J.R., Gallup G.G. Jr., Mirror self-recognition:
a review and critique of attempts to promote and engineer self-recognition in primates // Primates.
2015. V. 56. № 4. P. 317–326. doi 10.1007/s10329-0150488-9 18. Arshavsky Y.I. Cellular and network properties in the functioning of the nervous system: from central pattern generators to cognition // Brain Res. Rev. 2003.
V. 41 № 2–3. P. 229–267 19. Azevedo F.A.C., Carvalho L.R.B., Grinberg L.T., Farfel J.M., Ferretti R.E.L., Leite R.E.P., Filho W.J., Lent R., HerculanoHouzel S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain // J. Com. Neurol. 2009.
V. 513. № 5. P. 532–541. doi 10.1002/cne.21974 20. Baron-Cohen S., Leslie A.M., Frith U. Does the autistic child have a “theory of mind'' // Cognition. 1985;
21(1): 37–46.
21. Baron-Cohen S. Theory of mind and autism: a review.
Special Issue of the International Review of Mental Retardation. 2001. 23: 69–204 22. Berkeley G. An essay towards a new theory of vision.
Dublin: Pepyat; 1709.
23. Botvinick M., Jha A.P., Bylsma L.M., Fabian S.A., Solomon P.E., Prkachin K.M. Viewing facial expressions of pain engages cortical areas involved in the direct experience of pain // Neuroimage. 2005. V. 25. № 1. P. 312– 319. doi 10.1016/j.neuroimage.2004.11.043 24. Candidi M., Urgesi C., Ionta S., Aglioti S.M. Virtual lesion of ventral premotor cortex impairs visual perception of biomechanically possible but not impossible actions // Soc. Neurosci. 2008. V. 3 № 3–4. P. 388–400.
25. Carr L., Iacoboni M., Dubeau M.C., Mazziotta J.C., Lenzi G.L. Neural mechanisms of empathy in humans: a relay from neural systems for imitation to limbic areas // PNAS 2003. V. 100. № 9. P. 5497–5502 26. Carlezon W.A., Jr., Thomas M.J. Biological substrates of reward and aversion: a nucleus accumbens activity hypothesis // Neuropharmacol. 2009.V. 56. S. 1. P. 122–132.
27. Chartrand T.L., Bargh J.A. The chameleon effect: the perception-behavior link and social interaction // J.
Pers. Soc. Psychol. 1999. V. 76. № 6. P. 893–910.
doi 10.1037/0022-3514.76.6.893 28. Cheng Y.W., Tzeng O.J., Decety J., Imada T., Hsieh J.C.
Gender differences in the human mirror system: a magnetoencephalography study // Neuro Report. 2006. V. 17. № 11.
P. 1115–1119. doi 10.1097/01.wnr.0000223393.59328.21 29. Cheng Y., Decety J., Lin C.P., Hsieh J.C., Hung D., Tzeng O.J. Sex differences in spinal excitability during observation of bipedal locomotion // Neuro Report. 2007. V. 18. № 9. P. 887–890.
doi 10.1097/WNR.0b013e3280ebb486 30. Cheng Y., Yang C.Y., Lin C.P., Lee P.L., Decety J.
The perception of pain in others suppresses somatosensory oscillations: a magnetoencephalography study // Neuroimage. 2008. V. 40. № 4. P. 1833–1840.
doi 10.1016/j.neuroimage.2008.01.064 31. Cheng Y., Lee P.L., Yang C.Y., Lin C.P., Hung D., Decety J. Gender differences in the mu rhythm of the human mirror-neuron system // PLOS One. 2008. V. 3.
№ 5. e2113. doi 10.1371/journal.pone.0002113
32. Dapretto M., Davies M.S., Pfeifer J.H., Scott A.A., Sigman M., Bookheimer S.Y., Iacoboni M. Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders. // Nature Neuroscience.
2006. V. 9 № 1. P. 28–30. doi 10.1038/nn1611 33. Decety J. Naturaliser l’empathie [Empathy naturalized] // L’Encéphale. 2002. V. 28. № 1. P. 9–20.
34. Decety J., Jackson P.L The functional architecture of human empathy // Behav. Cog. Neurosc. Rev. 2004.
V. 3. № 2. P. 71–100. doi 10.1177/1534582304267187 35. Dinstein I., Thomas C., Behrmann M., Heeger D.J. A mirror up to nature. // Curr Biol. 2008. V. 18. № 1.
P. R13–R18. doi 10.1016/j.cub.2007.11.004 36. di Pellegrino G., Fadiga L., Fogassi L., Gallese V., Rizzolatti G. Understanding motor events: a neurophysiological study // Exp. Brain Res. 1992. V. 91. № 1.
P. 176–180.
37. Eichenbaum H. Memory on time // Trends Cogn. Sci.
2013. V. 17. № 2. P. 81–88.
38. Eichenbaum H. Time cells in the hippocampus: a new dimension for mapping memories // Nat. Rev. Neurosci.
2014. V. 15. № 11. P. 732–744.
39. Esber G.R., Holland P.C. The basolateral amygdala is necessary for negative prediction errors to enhance cue salience, but not to produce conditioned inhibition. // Eur. J. Neurosci. 2014. V. 40. № 9. P. 3328–3337.
40. Falck-Ytter T., Gredebäck G., von Hofsten C. Infants predict other people’s action goals. // Nat. Neurosci.
2006. V. 9. № 7. P. 878–879.
41. Fay W.H., Coleman R.O. A human sound transducer/ reproducer: Temporal capabilities of a profoundly echolalic child // Brain and Language. 1977. V. 4. № 3.
P. 396–402. doi 10.1016/0093-934X(77)90034-7 42. Fernando A.B.P., Murray J.E., Milton A.L. The amygdala:
securing pleasure and avoiding pain. // Front Behav Neurosci. 2013; 7: 190. Dec. 6, 2013.
doi 10.3389/fnbeh.2013.00190 43. Ferrari P.F., Gallese V., Rizzolatti G., Fogassi L. Mirror neurons responding to the observation of ingestive and communicative mouth actions in the ventral premotor cortex. // Eur. J. Neurosci. 2003. V. 17. № 8. P. 1703–1714.
44. Fogassi L., Ferrari P.F., Gesierich B., Rozzi S., Chersi F., Rizzolatti G. Parietal lobe: from action organization to intention understanding // Science. 2005. V. 308.
№5722. P. 662–667.
45. Freedberg D., Gallese V. Motion, emotion and empathy in esthetic experience // Trends Cogn. Scien. 2007.
V. 11. № 5. P. 197–203. doi 10.1016/j.tics.2007.02.003 46. Fried I., MacDonald K.A., Wilson C.L. Single neuron activity in human hippocampus and amygdala during recognition of faces and objects. // Neuron. 1997.
V. 18. № 5. P. 753–765.
47. Frith C.D., Frith U. Interacting minds: A biological basis // Science. 1999. V. 286. № 5445. P. 1692–1695 48. Gallese V., Fadiga L., Fogassi L., Rizzolatti G. Action recognition in the premotor cortex // Brain.1996.
V. 119. Pt. 2. P 593–609.
49. Gallese V. Agency and motor representations: new perspectives on intersubjectivity. ISC working papers 2000–2006. Workshop on autism and the theory of mind. Lyon, 8th May, 2000.
50. Gallese V., Goldman A. Mirror neurons and the simulation theory of mind-reading // Trends in Cogn. Scienc.
1998. V. 2. № 12. P. 493–501.
51. Gallese V. The “Shared Manifold” hypothesis: from mirror neurons to empathy // J. Consc. Stud. 2001.
V. 8. P. 33–50.
52. Gallese V., Fogassi L., Fadiga L., Rizzolati G. Action representation and the inferior parietal lobule. In:
Prinz W., Hommel B. editors. Attention & Performance XIX. Common Mechanisms in Perception and Action.
Oxford: Oxford University Press; 2002. P. 334–355 53. Gallese V. The roots of empathy: the shared manifold hypothesis and the neural basis of intersubjectivity // Psychopathology. 2003. V. 36. № 4. P. 171–180.
54. Gazzola V., Keysers C. The observation and execution of actions share motor and somatosensory voxels in all tested subjects: Single-subject analyses of unsmoothed fMRI data // Cereb. Cortex. 2009. V. 19 № 6. P. 1239–1255.
55. Goldman A. Interpretation psychologized // Mind and Language. 1989. V. 4. № 3. P 161–185. doi 10.1111/j.14680017.1989.tb00249.x 56. Gordon R. Folk psychology as simulation // Mind and Language. 1986. V. 1. № 2. P. 158–171. doi 10.1111/j.14680017.1986.tb00324.x 57. Hafting T., Fyhn M., Molden S., Moser M.B., Moser E.I.
Microstructure of a spatial map in the entorhinal cortex // Nature. 2005. V. 436. № 7052. P. 801–806.
58. Hennigan K., D’Ardenne K., McClure S.M. Distinct midbrain and habenula pathways are involved in processing aversive events in humans // J. Neurosci. 2015.
V. 35. № 1. P. 198–208.
59. Herculano-Houzel S. The remarkable, yet not extraordinary, human brain as a scaled-up primate brain and its associated cost // PNAS 2012. V. 109 Suppl. 1:
10661–10668. doi 10.1073/pnas.1201895109 60. Herculano-Houzel S., Avelino-de-Souza K., Neves K., Porfírio J., Messeder D., Mattos Feijó L., Maldonado J., Manger P.R. The elephant brain in numbers // Front Neuroanat. 2014. 8:46. doi 10.3389/fnana.2014.00046 61. Heyes C.M. Where do mirror neurons come from? // Neurosci Biobehav Rev. 2010. V. 34. № 4. P. 575–583.
62. Heyes C. Automatic imitation // Psycholog. Bul. 2011.
V 137. № 3. P. 463–483. doi 10.1037/a0022288 63. Hickok G. Eight problems for the mirror neuron theory of action understanding in monkeys and humans // J.
Cogn. Neurosci. 2009. V. 21. № 7. P. 1229–1243.
64. Jellema T., Baker C.I., Oram M.W., Perrett D.I. Cell populations in the banks of the superior temporal sulcus of the macaque monkey and imitation. In: Melzoff AN, Prinz W, editors. The imitative mind. Development, evolution and brain bases. Cambridge: Cambridge University Press; 2002. P. 267–290.
65. Kalat J.W. Biological Psychology. Belmont, CA: Wadsworth, Cengage Learning, 2009. 538–539.
66. Keysers C., Gazzola V. Integrating simulation and theory of mind: from self to social cognition // Trends in Cog. Scienc.
2007. V. 11. № 5. P. 194–196. doi 10.1016/j.tics.2007.02.002 67. Keysers C., Kaas J.H., Gazzola V. Somatosensation in social perception // Nat Rev Neurosci. 2010. V. 11.
№6. P. 417–428.
68. Kilner J.M., Neal A. Evidence of mirror neurons in human inferior frontal gyrus // J. Neurosci. 2009. V. 29. № 32, P. 10153–10159. doi 10.1523/jneurosci.2668-09.2009 69. Kropff E., Carmichael J.E., Moser M.B., Moser E.I.
Speed cells in the medial entorhinal cortex // Nature.
2015. V. 523. № 7561. P. 419–424.
70. Kosonogov V. Listening to action-related sentences impairs postural control // J. Electromyog. Kinesiol.
2011. V. 21. № 5. P. 742–745. doi 10.1016/j.jelekin.
2011.05.007 71. Kosonogov V. Why the mirror neurons cannot support action understanding // Neurophysiol. 2012. V. 44.
№6. P. 499–502. doi 10.1007/s11062-012-9327-4 72. Kreiman G., Koch C., Fried I. Category-specific visual responses of single neurons in the human medial temporal lobe // Nat Neurosci. 2000. V. 3. № 9. P. 946–953.
73. Iacoboni M. Understanding others: imitation, language, empathy. In: Perspectives on imitation: from cognitive neuroscience to social science, Hurley S., Chater N. (Eds), Cambridge, MA: MIT Press, 2005 74. Lakin J.L., Chartrand T.L. Using nonconscious behavioral mimicry to create affiliation and rapport // Psychological Science. 2003. V. 14. № 4. P. 334–339.
doi 10.1111/1467-9280.14481 75. Lamm C., Batson C.D., Decety J. The neural substrate of human empathy: effects of perspective-taking and cognitive appraisal // J. Cogn. Neurosc. 2007. V. 19.
№ 1. P. 42–58. doi 10.1162/jocn.2007.19.1.42 76. Liberman A.M., Cooper F.S., Shankweiler D.P., StuddertKennedy M. Perception of the speech code // Psychol.
Rev. 1967. V. 74. № 6. P. 431–461.
77. Lingnau A., Gesierich B., Caramazza A. Asymmetric fMRI adaptation reveals no evidence for mirror neurons in humans // PNAS. 2009. V. 106. № 24.
P. 9925–9930. doi 10.1073/pnas.0902262106 78. Longo M.R., Kosobud A., Bertenthal B.I. Automatic imitation of biomechanically possible and impossible actions: effects of priming movements versus goals // J. Exp. Psychol. Hum. Percept. Perform. 2008. V. 34.
№2. P. 489–501. doi 10.1037/0096-1523.34.2.489 79. Marslen-Wilson W. Linguistic structure and speech shadowing at very short latencies // Nature. 1973.
V. 244. № 5417. P. 522–523. doi 10.1038/244522a0 80. Mbemba J., Marte S., Christian K. Empathy for positive and negative emotions in the gustatory cortex // Neuroimage. 2007. V. 34. № 4. P. 1744–1753.
doi 10.1016/j.neuroimage.2006.10.032 81. McCarthy R., Warrington E.K. A two-route model of speech production. Evidence from aphasia // Brain: J.
Neurology. 1984. V. 107. № 2. P. 463–485.
doi 10.1093/brain/107.2.463 82. McCarthy R.A., Warrington E.K. Repeating without semantics:
surface dysphasia? // Neurocase. 2001. V. 7.
№ 1. P. 77–87. doi 10.1093/neucas/7.1.77 83. McRae K., Jones M.R. Semantic memory. The Oxford Handbook of Cognitive Psychology. Daniel R., ed.
New York, NY: Oxford University Press. 2013.
P. 206–216.
84. Medical Physiology. 2018. http://www.brainkart.
com/subject/Medical-Physiology_250/.
85. Meltzoff A.N., Decety J. What imitation tells us about social cognition: a rapprochement between developmental psychology and cognitive neuroscience // Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 2003. V. 358.
№ 1431. P. 491–500. doi 10.1098/rstb.2002.1261 86. Miller E.K., Cohen J.D. An integrative theory of prefrontal cortex function // Annu. Rev. Neurosci. 2001.
V. 24. P. 167–202.
87. Molenberghs P., Cunnington R., Mattingley J.B. Is the mirror neuron system involved in imitation? A short review and meta-analysis // Neurosc. Biobehav. Rev.
2009. V. 33 № 7. P. 975–980. doi 10.1016/j.neubiorev.
2009.03.010 88. Morrison I., Lloyd D., Di Pellegrino G., Roberts N. Vicarious responses to pain in anterior cingulate cortex:
Is empathy a multisensory issue? // Cogn. Affect. Behav.
Neurosc. 2004. V 4. № 2. P. 270–278.
doi 10.3758/CABN.4.2.270 89. Nambu A. Somatotopic organization of the primate basal ganglia // Front Neuroanat. 2011; 5: 26.
doi 10.3389/fnana.2011.00026 90. Nicoll R.A., Malenka R.C., Kauer J.A. Functional comparison of neurotransmitter receptor subtypes in mammalian central nervous system // Physiol. Rev.
1990. V. 70. № 2. P. 513–565.
91. Oberman L.M., Hubbard E.M., McCleery J.P., Altschuler E.L., Ramachandran V.S., Pineda J.A. EEG evidence for mirror neuron dysfunction in autism spectral disorders // Brain Res. Cogn. Brain Res.
2007. V. 24. № 2. P. 190–198.
92. Oberman L.M., Ramachandran V.S., Pineda J.A.
Modulation of mu suppression in children with autism spectrum disorders in response to familiar or unfamiliar stimuli: the mirror neuron hypothesis // Neuropsychologia. 2008. V. 46. № 5. P. 1558–65.
doi 10.1016/j.neuropsychologia.2008.01.010 93. Oberman L., Ramachandran V.S. Reflections on the mirror neuron system: Their evolutionary functions beyond motor representation. In Pineda, J.A. Mirror neuron systems: The role of mirroring processes in social cognition // Humana Press. 2009. P. 39–62.
94. O'Keefe J., Dostrovsky J. The hippocampus as a spatial map: preliminary evidence from unit activity in the freely moving rat // Brain Res. 1971. V. 31. № 1. 171–175.
95. O'Keefe J., Nadel L. The Hippocampus as a Cognitive Map. Oxford: Oxford U.P. 1978.
96. Pascolo P.B., Ragogna R., Rossi R. The mirror-neuron system paradigm and its consistency // Gait & Posture. 2009.
V. 30. Suppl. 1. 65. doi 10.1016/j.gaitpost.2009.07.064 97. Paukner A., Suomi S.J., Visalberghi E., Ferrari P.F. Capuchin monkeys display affiliation toward humans who imitate them // Science. 2009. V. 325. № 5942.
P. 880–883. doi 10.1126/science.1176269 98. Perkins T., Stokes M., McGillivray J., Bittar R. Mirror neuron dysfunction in autism spectrum disorders // J.
Clinic. Neurosc. 2010. V. 17. № 10. P. 1239–1243.
doi 10.1016/j.jocn.2010.01.026 99. Pillsbury WB. The place of movement in consciousness // Psychological Review. 1911. V. 18. № 2. P. 83–99.
100. Pineda J.A. Sensorimotor cortex as a critical component of an 'extended' mirror neuron system: Does it solve the development, correspondence, and control problems in mirroring? // Behav. Brain Funct. 2008;
4: 47. doi 10.1186/1744-9081-4-47 101. Petrides M., Cadoret G., Mackey S. Orofacial somatomotor responses in the macaque monkey homologue of Broca’s area // Nature. 2005. V. 435. № 7046.
P. 1235–1238.
102. Pobric G., Hamilton A.F.D c. Action understanding requires the left inferior frontal cortex // Current Biology. 2006. V. 16. № 5. P. 524–529.
doi 10.1016/j.cub.2006.01.033 103. Porter Jr.R.J., Lubker J.F. Rapid reproduction of vowel-vowel sequences: Evidence for a fast and direct acoustic-motoric linkage in speech // J.
Speech. Hear. Res. 1980. V. 23. № 3. P. 593–602.
doi 10.1044/jshr.2303.593 104. Preston S.D., de Waal F.B.M. Empathy: Its ultimate and proximate bases // Behavioral and Brain Sciences. 2002.
V. 25. № 1. P. 1–20. doi 10.1017/s0140525x02000018 105. Quiroga R.Q., Reddy L., Kreiman G., Koch C., Fried I.
Invariant visual representation by single neurons in the human brain // Nature. 2005. V. 435. № 7045. P. 1102–1107.
106. Ramachandran V.S. Self awareness: the last frontier, Edge Foundation web essay. (2009) Retrieved July 26, 2011.
107. Redolar-Ripoll D., Soriano-Mas C., Guillazo-Blanch G., Aldavert-Vera L., Segura-Torres P., Morgado-Bernal I.
Posttraining intracranial self-stimulation ameliorates the detrimental effects of parafascicular thalamic lesions on active avoidance in young and aged rats // Behav.
Neurosci. 2003. V. 117. № 2. P. 246–256.
108. Reniers R.L., Beavan A., Keogan L., Furneaux A., Mayhew S., Wood S.J. Is it all in the reward? Peers influence risk-taking behaviour in young adulthood // Br.
J. Psychol. 2017 V. 108. № 2. P. 276–295. [Epub ahead of print] doi 10.1111/bjop.12195 109. Rizzolatti G., Craighero L. The Mirror-Neuron System // Annu. Rev. Neurosci. 2004. V 27. P. 169–192.
110. Rizzolatti G., Fadiga L., Gallese V., Fogassi L. Premotor cortex and the recognition of motor actions // Cogn.
Brain Res. 1996, V. 3. № 2. P. 131–141.
111. Rizzolatti G., Fadiga L., Fogassi L., Gallese V. Resonance behaviors and mirror neurons. // Arch. Ital. Biol.
1999. V. 137. № 2–3. P. 85–100.
112. Rizzolatti G., Fogassi L., Gallese V. Neurophysiological mechanisms underlying the understanding and imitation of action // Nat. Rev. Neurosci. 2001. V. 2. № 9.
P. 661–670.
113. Rizzolatti G., Craighero L. The Mirror-Neuron System // Annu. Rev. Neurosci. 2004. V. 27. P. 169–192.
114. Ruiz-Medina J., Redolar-Ripoll D., Morgado-Bernal I., Aldavert-Vera L., Segura-Torres P. Intracranial selfstimulation improves memory consolidation in rats with little training // Neurobiol. Learn. Mem. 2008.
V. 89. № 4. P. 574–581.
115. Sargolini F., Fyhn M., Hafting T., McNaughton B.L., Witter M.P., Moser M.B., Moser E.I. Conjunctive representation of position, direction, and velocity in entorhinal cortex // Science. 2006. V. 312 № 5774. P. 758–762.
116. Saygin A.P. Superior temporal and premotor brain areas necessary for biological motion perception // Brain: J. Neurology. 2007. V. 130. Pt. 9. P. 2452–2461.
doi 10.1093/brain/awm162 117. Saygin A.P., Wilson S.M., Dronkers N.F., Bates E. Action comprehension in aphasia: Linguistic and nonlinguistic deficits and their lesion correlates // Neuropsychologia.
2004. V. 42. № 13. P.1788–1804.
118. Scheerer E. Motor theories of cognitive structure: A historical review. In: Prinz W, Sanders AF, editors.
Cognition and Motor Processes // Berlin: SpringerVerlag. 1984. P. 77–98.
119. Schippers M.B., Roebroeck A., Renken R., Nanetti L., Keysers C. Mapping the Information flow from one brain to another during gestural communication // Proc. Natl. Acad. Sci. USA. 2010. V. 107. № 20.
P. 9388–9393. doi 10.1073/pnas.1001791107 120. Schulte-Rüther M., Markowitsch H.J., Shah N.J., Fink G.R., Piefke M. Gender differences in brain networks supporting empathy // Neuroimage. 2008. V. 42. № 1.
P. 393–403. doi 10.1073/pnas.100179110710.1016/j.neuroimage.
2008.04.180 121. Schultz W. Predictive reward signal of dopamine neurons // J. Neurophysiol. 1998. V. 80. № 1. P. 1–27.
122. Schultz W., Dickinson A. Neuronal coding of prediction errors // Annu. Rev. Neurosci. 2000. V. 23. 473–500.
123. Segura-Torres P., Capdevila-Ortнs L., Martн-Nicolovius M., Morgado-Bernal I. Improvement of shuttle-box learning with pre- and post-trial intracranial selfstimulation in rats // Behav. Brain Res. 1988. V. 29. № 1–2.
P. 111–117.
124. Segura-Torres P., Portell-Cortйs I., Morgado-Bernal I.
Improvement of shuttlebox avoidance with post-training intracranial self-stimulation, in rats: a parametric study // Behav. Brain Res. 1991. V. 42. № 2. P. 161–167.
125. Sommerville J.A., Decety J. Weaving the fabric of social interaction: articulating developmental psychology and cognitive neuroscience in the domain of motor cognition // Psychonom. Bulletin, Review. 2006.
V. 13. № 2. P. 179–200. doi 10.3758/BF03193831 126. Sun C., Kitamura T., Yamamoto J., Martin J., Pignatelli M., Kitch L.J., Schnitzer M.J., Tonegawa S. Distinct speed dependence of entorhinal island and ocean cells, including respective grid cells // PNAS USA. 2015.
V. 112. № 30. P. 9466–9471.
127. Tanji J., Hoshi E. Role of the lateral prefrontal cortex in executive behavioral control // Physiol. Rev. 2008.
V. 88. № 1. P. 37–57.
128. Tkach D., Reimer J., Hatsopoulos N.G. Congruent activity during action and action observation in motor cortex // J. Neurosci. 2007. V. 27. № 48. P. 13241–13250.
129. Thompson L.T., Best P.J. Long-term stability of the place-field activity of single units recorded from the dorsal hippocampus of freely behaving rats // Brain Res. 1990. V. 509. № 2. P. 299–308.
130. Tobler P.N., Dickinson A., Schultz W. Coding of predicted reward omission by dopamine neurons in a conditioned inhibition paradigm // J. Neurosci. 2003.
V. 23. № 32. P. 10402–10410.
131. Tolman E.C. Cognitive maps in rats and men // Psychol.
Rev. 1948. V. 55 № 4. P. 189–208.
132. Tomita H., Ohbayashi M., Nakahara K., Hasegawa I., Miyashita Y. Top-down signal from prefrontal cortex in executive control of memory retrieval // Nature.
1999. V. 401. № 6754. P. 699–703.
133. Tower D.B. Structural and functional organization of mammalian cerebral cortex; the correlation of neuron density with brain size; cortical neuron density in the fin whale (Balaenoptera physalus L.) with a note on the cortical neurone density in the Indian elephant // J. Comp. Neurol. 1954. V. 101. № 1. P. 19–51.
doi 10.1002/cne.901010103 134. Tranel D., Kemmerer D., Adolphs R., Damasio H., Damasio A.R. Neural correlates of conceptual knowledge for actions // Cogn. Neuropsychol. 2003. V. 20.
№ 3. P. 409–432. doi 10.1080/02643290244000248 135. Tulving E. Episodic and semantic memory. In Organization of Memory, ed. E Tulving, W Donaldson, New York: Academic. 1972. P. 381–403.
136. Ullman M.T. Contributions of memory circuits to language:
the declarative/procedural model // Cognition 2004. V. 92. № 1–2. P. 231–270.
137. van Baaren R.B., Maddux W.W., Chartrand T.L., De Bouter C., van Knippenberg A. It takes two to mimic:
behavioral consequences of self-construals // J. Personal.
Social Psychology. 2003. V. 84. № 5. P. 1093–1102.
doi 10.1037/0022-3514.84.5.1093 138. Washburn M.F. The function of incipient motor processes // Psychological Review. 1914. V. 21. № 5.
P. 376–390.
139. Washburn M.F. Movement and mental imagery: Outlines of a motor theory of the complexer mental processes // Boston: Houghton Mifflin; 1916.
140. Watson J.B. Psychology as the behaviorist views it // Psychological Review. 1913. V. 20 № 2. P. 158–177.
141. Wicker B., Keysers C., Plailly J., Royet J.P., Gallese V., Rizzolatti G. Both of us disgusted in My insula: the common neural basis of seeing and feeling disgust // Neuron. 2003. V. 40. № 3. P. 655–664.
142. Wu M., Kujawa A., Lu L.H., Fitzgerald D.A., Klumpp H., Fitzgerald K.D., Monk C.S., Phan K.L. Age-related changes in amygdala-frontal connectivity during emotional face processing from childhood into young adulthood // Hum Brain Mapp. 2016. V. 37. № 5.
P. 1684–1695. [Epub ahead of print] doi 10.1002/hbm.23129 143. Yang C.Y., Decety J., Lee S., Chen C., Cheng Y. Gender differences in the mu rhythm during empathy for pain: an electroencephalographic study // Brain Res. 2009.
V. 1251. P. 176–184. doi 10.1016/j.brainres.2008.11.062
Источник:
Дата создания: 05.03.2024
Последнее редактирование: 11.03.2024
Относится к аксиоматике: Системная нейрофизиология.
Другие страницы раздела "Отзеркаливание чужих навыков":
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя